Understory Vegetation Change Following Woodland Reduction Varies by Plant Community Type and Seeding Status: A Region-Wide Assessment of Ecological Benefits and Risks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
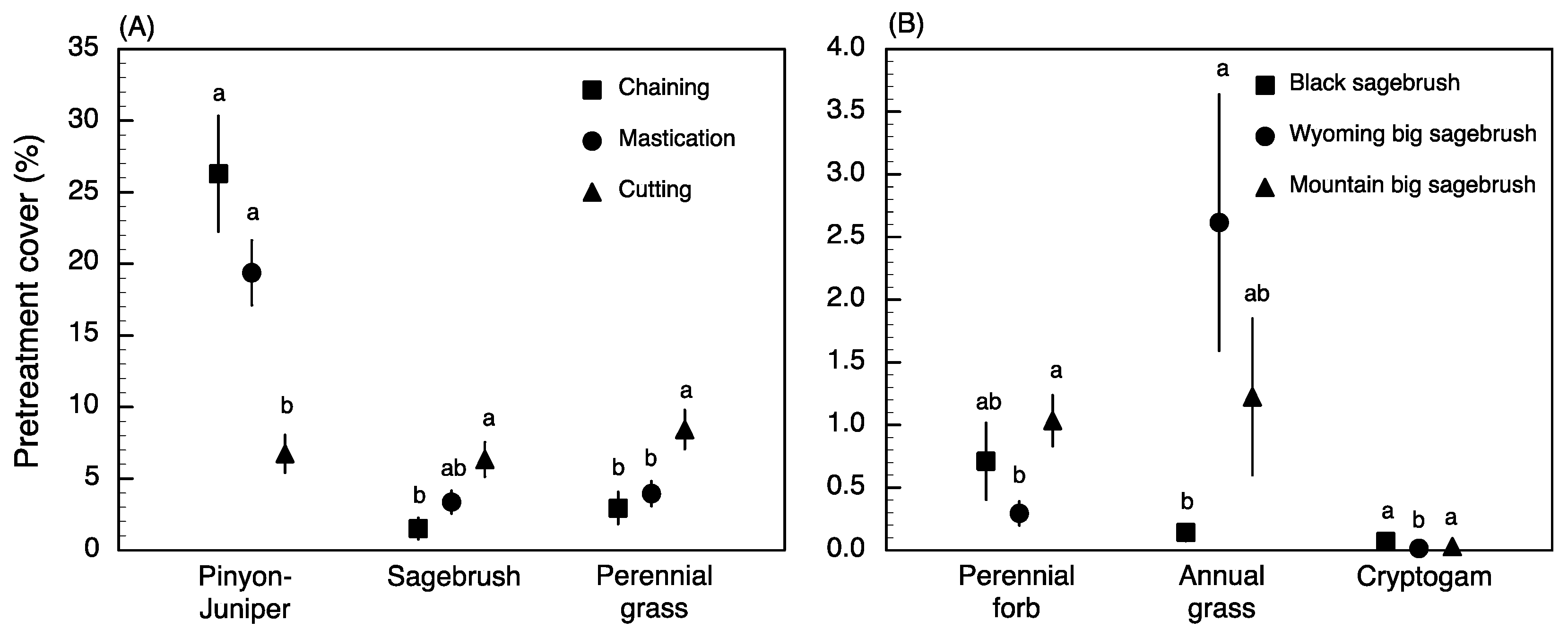
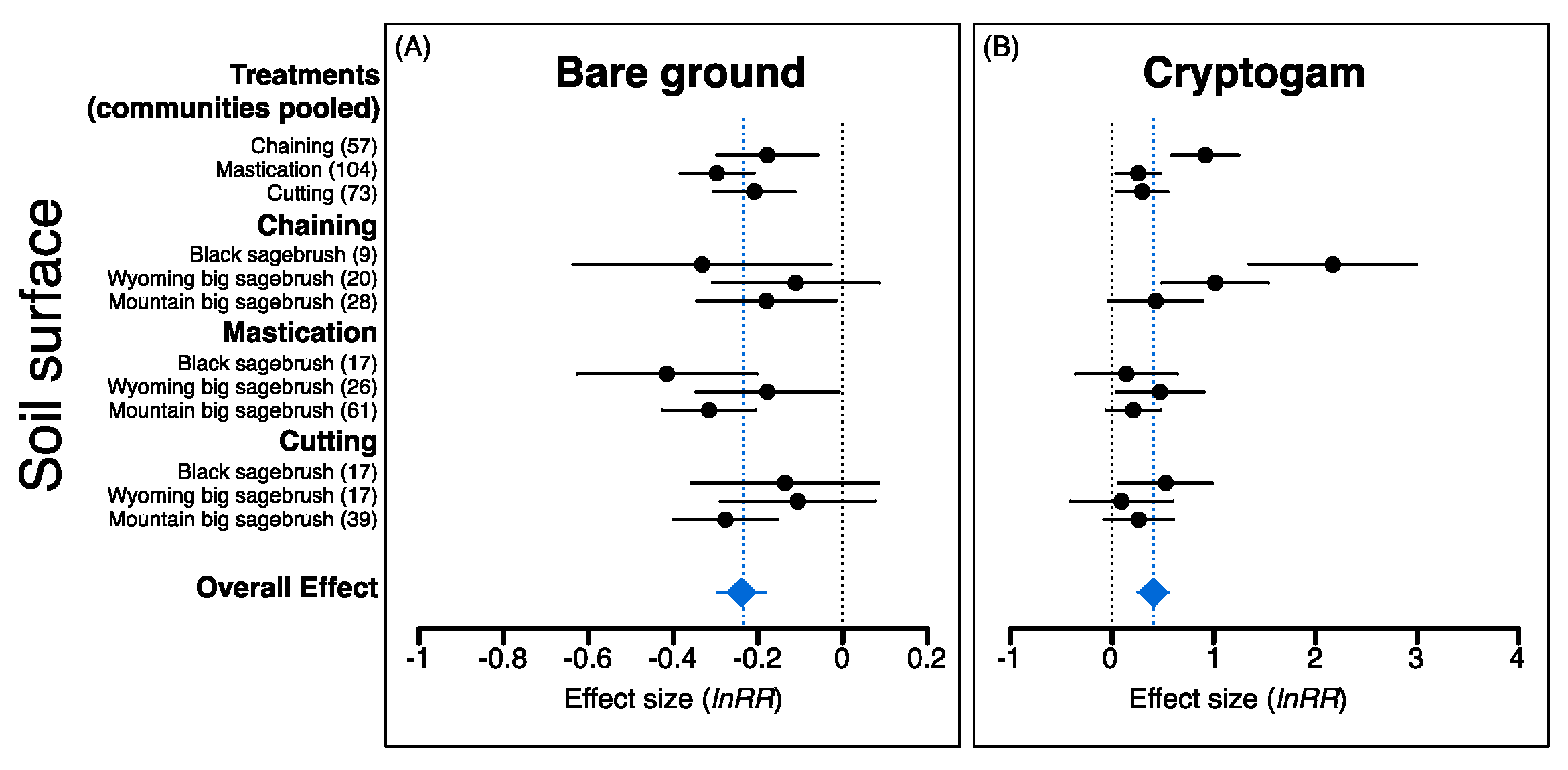
2.1. Pre-Treatment Vegetation
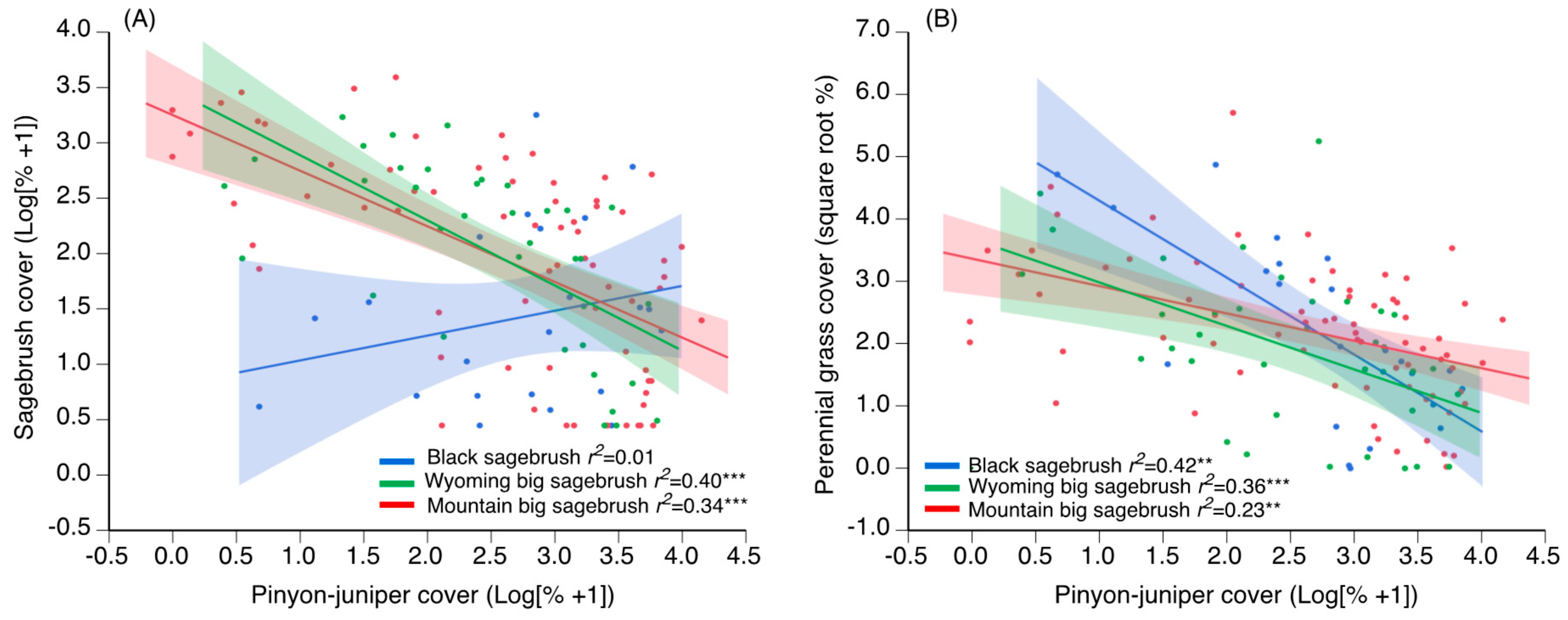
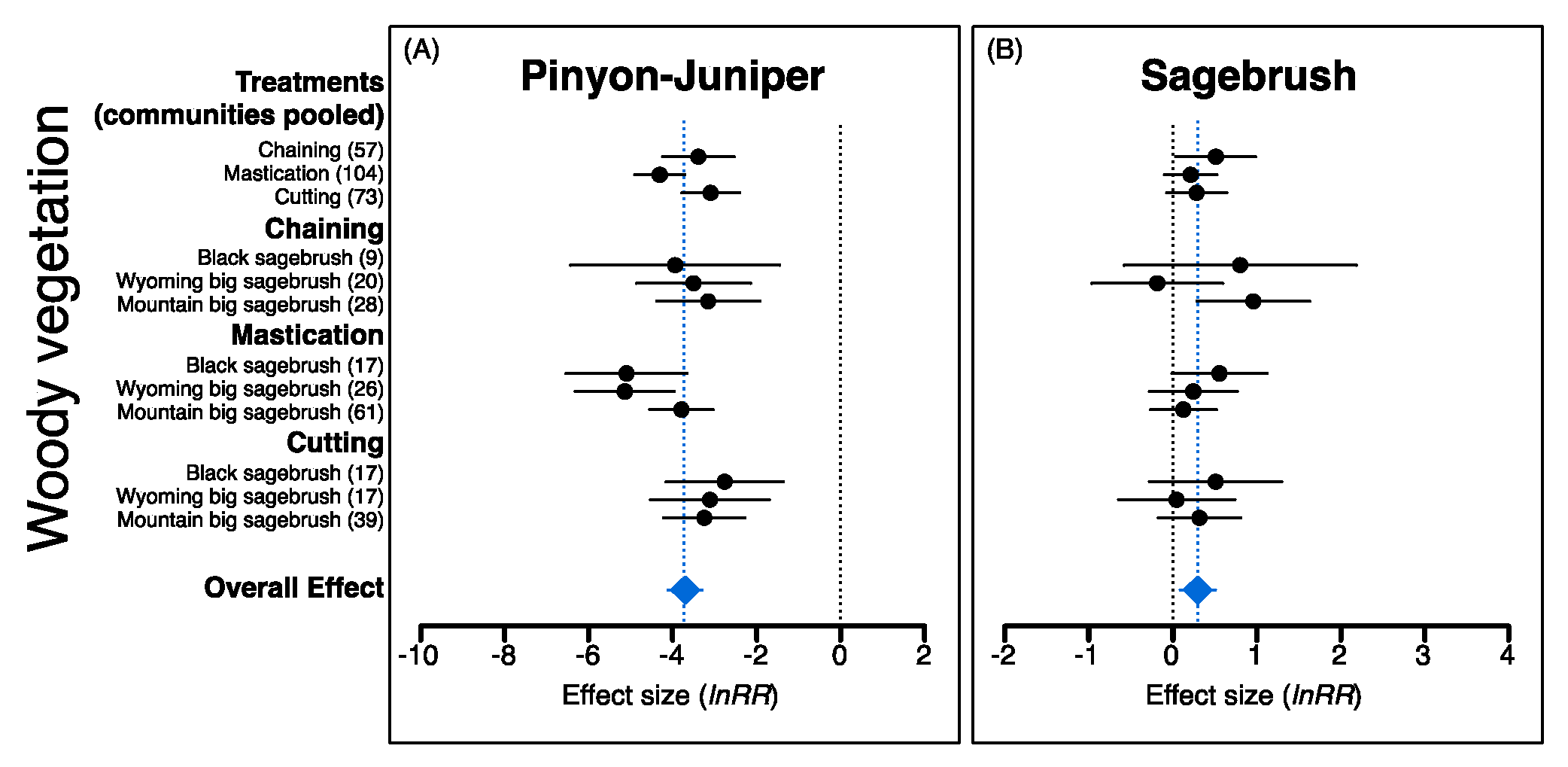
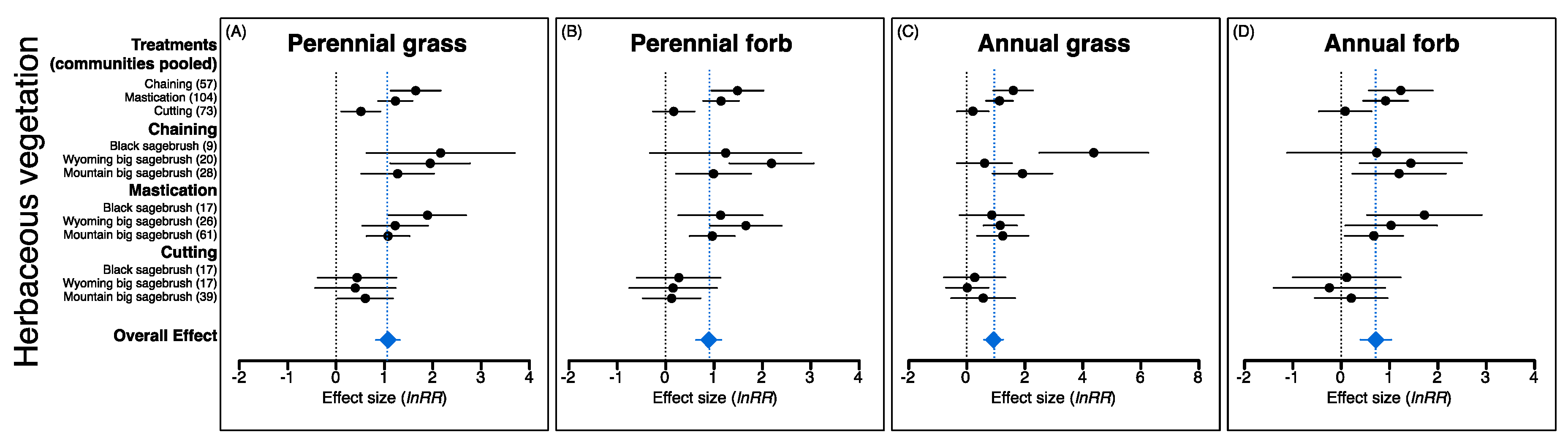
2.2. Plant Community Responses to P–J Reduction
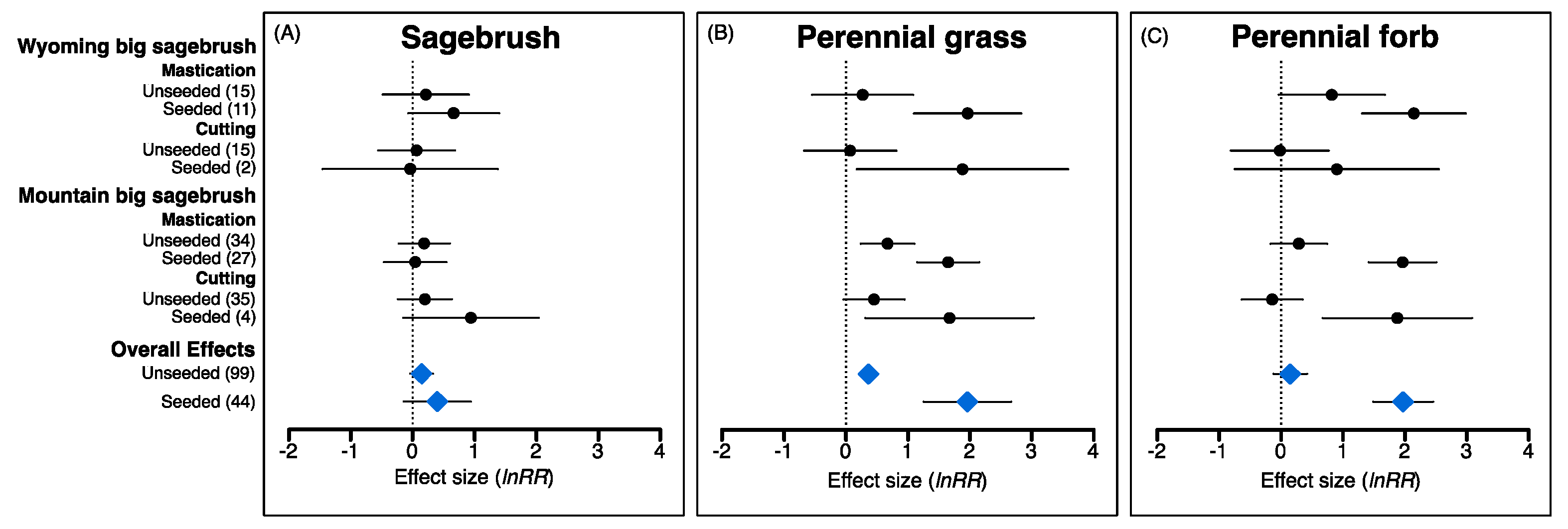
2.3. Influence of Seeding on Post-Treatment Understory Recovery
3. Discussion
3.1. Overall Effects of P–J Reduction and Seeding were Dependent on Disturbance Intensity and Pre-Treatment Vegetation
3.2. Why Were Understory Responses More Pronounced for Chaining and Mastication Than Cutting?
3.3. What Ecological Processes Were Responsible for the Differences in Understory Resilience and Annual Grass Response among Plant Community Types?
3.4. When Is Seeding Essential for Understory Recovery Following P–J Reduction?
3.5. Conclusions
4. Materials and Methods
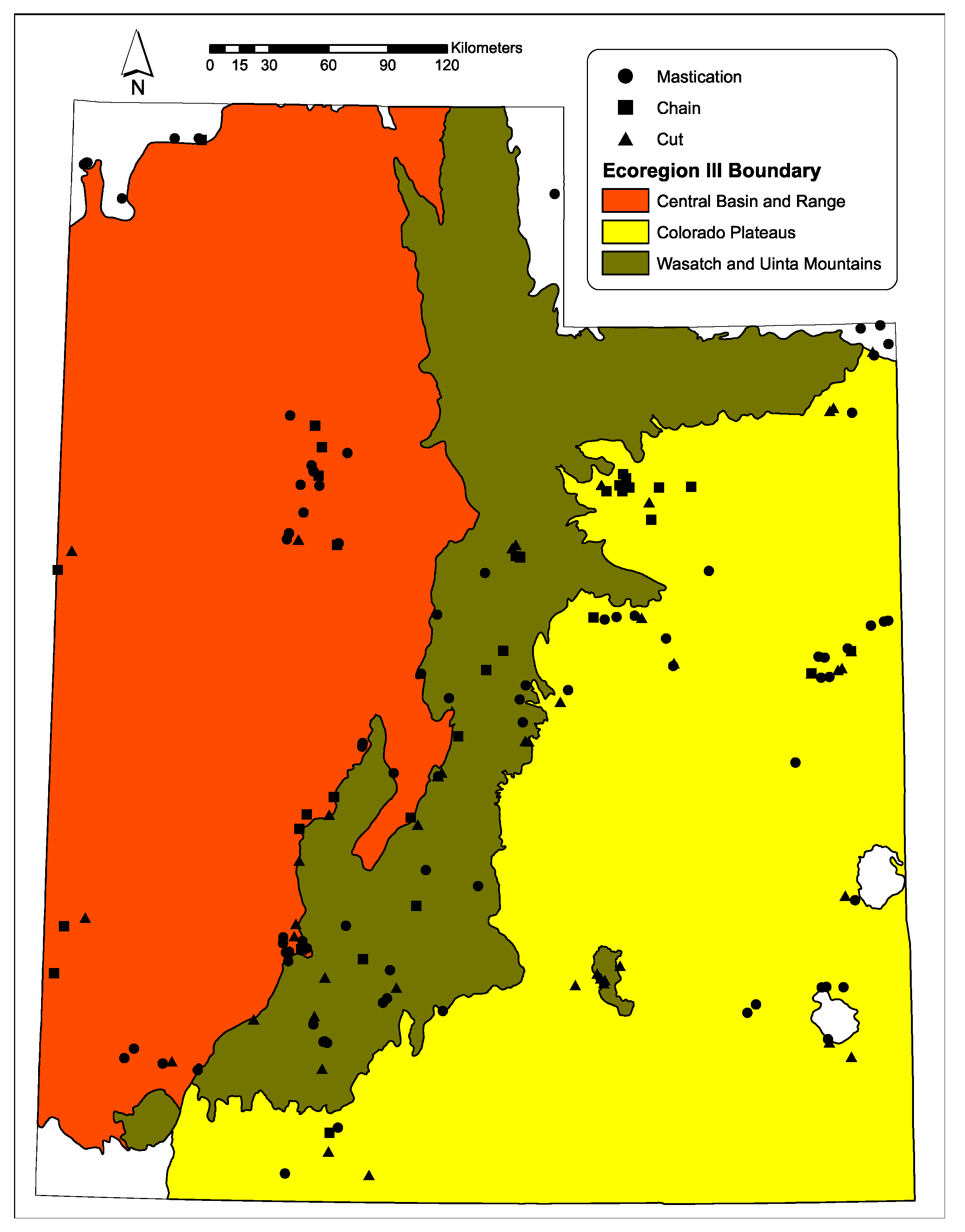
4.1. Project Sites
4.2. Treatments and Seeding
4.3. Vegetation and Ground Surface Sampling
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilcox, B.P.; Birt, A.; Fuhlendorf, S.D.; Archer, S.R. Emerging frameworks for understanding and mitigating woody plant encroachment in grassy biomes. Curr. Opin. Environ. Sustain. 2018, 32, 46–52. [Google Scholar] [CrossRef]
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody plant encroachment: Causes and consequences. In Rangeland Systems: Processes, Management and Challenges; Briske, D.D., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 25–83. [Google Scholar]
- Romme, W.H.; Allen, C.D.; Balley, J.D.; Baker, W.L.; Bestelmeyer, B.T.; Brown, P.M.; Eisenhart, K.S.; Floyd, M.L.; Huffman, D.W.; Jacobs, B.F.; et al. Historical and modern disturbance regimes, stand structures, and landscape dynamics in pinon-juniper vegetation of the western United States. Rangel. Ecol. Manag. 2009, 62, 203–222. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.; Svejcar, T.; Rose, J. Conversion of shrub steppe to juniper woodland. In Proceedings: Ecology and Mangement of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; Proc. RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 385–390. [Google Scholar]
- Miller, R.F.; Chambers, J.C.; Evers, L.; Williams, C.J.; Snyder, K.A.; Roundy, B.A.; Pierson, F.B. The Ecology, History, Ecohydrology, and Management of Pinyon Juniper Woodlands in the Great Basin and Northern Colorodao Plateau of the Western United States; Gen. Tech. Rep. RMRS-GTR-403; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2019; p. 284.
- Swetnam, T.W.; Allen, C.D.; Betancourt, J.L. Applied historical ecology: Using the past to manage the future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Baker, W.L.; Shinneman, D.J. Fire and restoration of pinon-juniper woodlands in the western United States: A review. For. Ecol. Manag. 2004, 189, 1–21. [Google Scholar] [CrossRef]
- Burkhardt, J.W.; Tisdale, E.W. Causes of juniper invasion in southwestern Idaho. Ecology 1976, 57, 472–484. [Google Scholar] [CrossRef]
- Bauer, J.M.; Weisberg, P.J. Fire history of a central Nevada pinyon-juniper woodland. Can. J. For. Res. 2009, 39, 1589–1599. [Google Scholar] [CrossRef]
- Shinneman, D.J.; Baker, W.L. Historical fire and multidecadal drought as context for pinon-juniper woodland restoration in western Colorado. Ecol. Appl. 2009, 19, 1231–1245. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.F.; Rose, J.A. Fire history and western juniper encroachment in sagebrush steppe. J. Range Manag. 1999, 52, 550–559. [Google Scholar] [CrossRef]
- Miller, R.F.; Tausch, R.J.; McArthur, E.D.; Johnson, D.D.; Sanderson, S.C. Age Structure and Expansion of Piñon-Juniper Woodlands: A Regional Perspective in the Intermountain West; RMRS-RP-69; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ed.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; p. 17.
- Miller, R.E.; Rose, J.A. Historic expansion of Juniperus occidentalis (western juniper) in Southeastern Oregon. Great Basin Nat. 1995, 55, 37–45. [Google Scholar]
- Blackburn, W.H.; Tueller, P.T. Pinyon and juniper invasion in black sagebrush communities in east-central Nevada. Ecology 1970, 51, 841–848. [Google Scholar] [CrossRef]
- Mollnau, C.; Newton, M.; Stringham, T. Soil water dynamics and water use in a western juniper (Juniperus occidentalis) woodland. J. Arid Environ. 2014, 102, 117–126. [Google Scholar] [CrossRef]
- Miller, R.F.; Wigand, P.E. Holocene changes in semiarid pinyon-juniper woodlands. BioScience 1994, 44, 465–474. [Google Scholar] [CrossRef]
- Miller, R.F.; Tausch, R.J. The role of fire in juniper and pinyon woodlands: A descriptive analysis. In Fire Conference 2000: The first National Congress on Fire Ecology, Prevention, and Management, Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species, 27 November–1 December 2000, San Diego, CA; Misc. Pub. No. 11; Galley, K.E.M., Wilson, T.P., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2001; pp. 15–30. [Google Scholar]
- Fritts, H.C. Relationships of ring widths in arid- site conifers to variations in monthly temperature and precipitation. Ecol. Monogr. 1974, 44, 411–440. [Google Scholar] [CrossRef]
- Miller, R.F.; Knick, S.T.; Pyke, D.A.; Meinke, C.W.; Hanser, S.E.; Wisdom, M.J.; Hild, A.L. Characteristics of sagebrush habitats and limitations to long-term conservation. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and its Habitats; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 145–184. [Google Scholar]
- Cottam, W.P.; Stewart, G. Plant succession as a result of grazing and of meadow dessication by erosion since settlement in 1862. J. For. 1940, 38, 613–626. [Google Scholar]
- Ko, D.W.; Sparrow, A.D.; Weisberg, P.J. Land-use legacy of historical tree harvesting for charcoal production in a semi-arid woodland. For. Ecol. Manag. 2011, 261, 1283–1292. [Google Scholar] [CrossRef]
- West, N.E. Intermountain deserts, shrub steppes, and woodlands. In North American Terrestrial Vegetation; Barbour, M.G., Billings, W.D., Eds.; Cambridge University Press: New York, NY, USA, 1988; pp. 209–230. [Google Scholar]
- Mitchell, J.E.; Roberts, T.C. Distribution of pinyon-juniper in the western United States. In Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 146–154. [Google Scholar]
- Tueller, P.T.; Beeson, C.D.; Tausch, R.J.; West, N.E.; Rea, K.H. Pinyon-Juniper Woodlands of the Great Basin: Distribution, Flora, Vegetatal Cover; Res. Rep. INT-229; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1979; p. 22.
- Evans, R.A. Management of Pinyon-Juniper Woodlands; Gen. Tech. Rep. INT-249; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1988; p. 34.
- (GLTI). Inventorying, Classifying, and Correlating Juniper and Pinyon Communities to Soils in Western United States; U.S. Department of Agriculture, Natural Resources Conservation Service, Grazing Lands Technology Institute: Fort Worth, TX, USA, 1997; p. 39.
- Krueger, W.C.; Winward, A.H. Influence of cattle and big game grazing on understory structure of a douglas fir-ponderosa pine-kentucky bluegrass community. J. Range Manag. 1974, 27, 450–453. [Google Scholar] [CrossRef]
- Weisberg, P.J.; Lingua, E.; Pillai, R.B. Spatial patterns of pinyon-juniper woodland expansion in central Nevada. Rangel. Ecol. Manag. 2007, 60, 115–124. [Google Scholar] [CrossRef]
- Page, D.; Gottfried, G.; Tausch, R.; Lanner, R.; Ritter, S. Management of Pinyon-Juniper “Woodland” Ecosystems. A Position of the Intermountain Society of American Foresters. 2013. Available online: http://www.usu.edu/saf/PJWoodlandsPositionStatement.pdf (accessed on 27 August 2020).
- Tausch, R.J.; West, N.E.; Nabi, A.A. Tree age and dominance patterns in Great-Basin pinyon-juniper woodlands. J. Range Manag. 1981, 34, 259–264. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef]
- Davies, K.W.; Boyd, C.S.; Beck, J.L.; Bates, J.D.; Svejcar, T.J.; Gregg, M.A. Saving the sagebrush sea: An ecosystem conservation plan for big sagebrush plant communities. Biol. Conserv. 2011, 144, 2573–2584. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.F.; Naugle, D.E.; Maestas, J.D.; Hagen, C.A.; Hall, G. Special issue: Targeted woodland removal to recover at-risk grouse and their sagebrush-steppe and prairie ecosystems. Rangel. Ecol. Manag. 2017, 70, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Petersen, S.L.; Stringham, T.K. Intercanopy community structure across a heterogeneous landscape in a western juniper-encroached ecosystem. J. Veg. Sci. 2009, 20, 1163–1175. [Google Scholar] [CrossRef]
- Burkhardt, J.W.; Tisdale, E.W. Nature and successional status of western juniper in Idaho. J. Range Manag. 1969, 22, 264–270. [Google Scholar] [CrossRef]
- West, N.E. Basic synecological relationshiops of sagebrush-dominated lands in the Great Basin and the Colorado Plateau. In The Sagebrush Ecosystem: A Symposium; Utah State University, College of Natural Resources: Logan, UT, USA, 1979; pp. 33–41. [Google Scholar]
- Tausch, R.J.; Tueller, P.T. Foliage biomass and cover relationships between tree-dominated and shrub-dominated communities in pinyon-juniper woodlands. Great Basin Nat. 1990, 50, 121–134. [Google Scholar]
- Tausch, R.J.; West, N.E. Plant species composition patterns with differnces in tree dominance on a southwestern Utah pinyon-juniper site. In Desired Future Conditions for Pinyon-Juniper Ecosystems, 8–12 August 1994, Flagstaff, AZ; Gen. Tech. Rep. RM-258; Shaw, D.W., Aldon, E.F., LoSapio, C., Eds.; U.S. Department of Agriculture, Rocky Mountain Forest and Range Experimental Station: Fort Collins, CO, USA, 1995; pp. 16–23. [Google Scholar]
- Roundy, B.A.; Miller, R.F.; Tausch, R.J.; Young, K.; Hulet, A.; Rau, B.; Jessop, B.; Chambers, J.C.; Eggett, D. Understory cover responses to piñon–juniper treatments across tree dominance gradients in the Great Basin. Rangel. Ecol. Manag. 2014, 67, 482–494. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, T.E.; Haufler, J.B.; Stelter, L.H.; Nagy, J.G. Nongame wildlife responses to chaining of pinyon-juniper woodlands. J. Wildl. Manag. 1981, 45, 381–389. [Google Scholar] [CrossRef]
- Rau, B.M.; Tausch, R.; Reiner, A.; Johnson, D.W.; Chambers, J.C.; Blank, R.R. Developing a model framework for predicting effects of woody expansion and fire on ecosystem carbon and nitrogen in a pinyon-juniper woodland. J. Arid Environ. 2012, 76, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Short, H.L.; Evans, W.; Boeker, E.L. The use of natural and modified pinyon pine Juniper woodlands by deer and elk. J. Wildl. Manag. 1977, 41, 543–559. [Google Scholar] [CrossRef]
- Bristow, N.A.; Weisberg, P.J.; Tausch, R.J. A 40-year record of tree establishment following chaining and prescribed fire treatments in singleleaf pinyon (Pinus monophylla) and Utah juniper (Juniperus osteosperma) Woodlands. Rangel. Ecol. Manag. 2014, 67, 389–396. [Google Scholar] [CrossRef]
- Bates, J.D.; Davies, K.W. Effects of conifer treatments on soil nutrient availability and plant composition in sagebrush steppe. For. Ecol. Manag. 2017, 400, 631–644. [Google Scholar] [CrossRef]
- Jameson, D.A. The relationship of tree overstory and herbaceous understory vegetation. J. Range Manag. 1967, 20, 247–249. [Google Scholar] [CrossRef]
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.Y.; Morton, J.A.; Knapp, A.K. Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance. J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Ann. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Roundy, B.A.; Farmer, M.; Olson, J.; Petersen, S.; Nelson, D.R.; Davis, J.; Vernon, J. Runoff and sediment response to tree control and seeding on a high soil erosion potential site in Utah: Evidence for reversal of an abiotic threshold. Ecohydrology 2017, 10, 1–9. [Google Scholar] [CrossRef]
- Miller, R.F.; Svejcar, T.J.; Rose, J.A. Impacts of western juniper on plant community composition and structure. J. Range Manag. 2000, 53, 574–585. [Google Scholar] [CrossRef]
- Pierson, F.B.; Williams, C.J.; Kormos, P.R.; Hardegree, S.P.; Clark, P.E.; Rau, B.M. Hydrologic vulnerability of sagebrush steppe following pinyon and juniper encroachment. Rangel. Ecol. Manag. 2010, 63, 614–629. [Google Scholar] [CrossRef]
- Huxman, T.E.; Wilcox, B.P.; Breshears, D.D.; Scott, R.L.; Snyder, K.A.; Small, E.E.; Hultine, K.; Pockman, W.T.; Jackson, R.B. Ecohydrological implications of woody plant encroachment. Ecology 2005, 86, 308–319. [Google Scholar] [CrossRef]
- Williams, C.J.; Pierson, F.B.; Al-Hamdan, O.Z.; Kormos, P.R.; Hardegree, S.P.; Clark, P.E. Can wildfire serve as an ecohydrologic threshold-reversal mechanism on juniper-encroached shrublands? Ecohydrology 2014, 7, 453–477. [Google Scholar] [CrossRef]
- Madsen, M.D.; Zvirzdin, D.L.; Petersen, S.L.; Hopkins, B.G.; Roundy, B.A.; Chandler, D.G. Soil water repellency within a burned pinon-juniper woodland: Spatial distribution, severity, and ecohydrologic implications. Soil Sci. Soc. Am. J. 2011, 75, 1543–1553. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, C.L.; Walker, S.C.; Stevens, R. Soil seed banking in pinyon-juniper areas with differing levels of tree cover, understory density and composition. In Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; Proc. RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 141–145. [Google Scholar]
- Naillon, D.; Memmott, K.; Monsen, S.B. A comparison of understory species at three densities in a pinyon-juniper woodland. In Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; Proc. RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, Utah, USA, 1999; pp. 72–75. [Google Scholar]
- Koniak, S.; Everett, R.L. Seed reserves in soils of successional stages of pinyon woodlands. Am. Midl. Nat. 1982, 108, 295–303. [Google Scholar] [CrossRef]
- Allen, E.A.; Nowak, R.S. Effect of pinyon–juniper tree cover on the soil seed bank. Rangel. Ecol. Manag. 2008, 61, 63–73. [Google Scholar] [CrossRef]
- Suminski, R.R. Management implications for mule dear winter range in northern pinyon-juniper. In Managing Pinyon-Juniper Ecosystems for Sustainability and Social Need, 26–30 April 1993, Sante Fe, NM; Gen Tech. Rep. RM-236; Aldon, E.F., Shaw, D.W., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Moutain Forest and Range Experiment Staion: Fort Collins, CO, USA, 1993; pp. 133–139. [Google Scholar]
- Commons, M.L.; Baydack, R.K.; Braun, C.E. Sage grouse response to pinyon-juniper management. In Proceedings: Ecology and Management of Pinyon-Juniper Communities in the Interior West, 15–18 September 1997, Provo, UT; RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 238–239. [Google Scholar]
- Frey, S.N.; Curtis, R.; Heaton, K. Response of a small population of greater sage-grouse to tree removal: Implications of limiting factors. Hum. Wildl. Interact. 2013, 7, 260–272. [Google Scholar]
- Baruch-Mordo, S.; Evans, J.S.; Severson, J.P.; Naugle, D.E.; Maestas, J.D.; Kiesecker, J.M.; Falkowski, M.J.; Hagen, C.A.; Reese, K.P. Saving sage-grouse from the trees: A proactive solution to reducing a key threat to a candidate species. Biol. Conserv. 2013, 167, 233–241. [Google Scholar] [CrossRef]
- Hagen, C.A. Greater Sage-Grouse Conservation Assessment and Strategy for Oregon: A Plan To Maintain and Enhance Populations of Habitat; Oregon Department of Fish and Wildlife: Salem, OR, USA, 2011; p. 207. [Google Scholar]
- Knick, S.T.; Hanser, S.E.; Leu, M. Ecological scale of bird community response to piñon-juniper removal. Rangel. Ecol. Manag. 2014, 67, 553–562. [Google Scholar] [CrossRef]
- Crawford, J.A.; Olson, R.A.; West, N.E.; Mosley, J.C.; Schroeder, M.A.; Whitson, T.D.; Miller, R.F.; Gregg, M.A.; Boyd, C.S. Ecology and management of sage-grouse and sage-grouse habitat. J. Range Manag. 2004, 57, 2–19. [Google Scholar] [CrossRef] [Green Version]
- Tausch, R.J. Transitions and thresholds: Influences and implications for management in pinyon and juniper woodlands. In Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 361–365. [Google Scholar]
- Davies, K.W.; Bates, J.D.; Boyd, C.S. Postwildfire seeding to restore native vegetation and limit exotic annuals: An evaluation in juniper-dominated sagebrush steppe. Restor. Ecol. 2019, 27, 120–127. [Google Scholar] [CrossRef]
- Miller, R.F.; Ratchford, J.; Roundy, B.A.; Tausch, R.J.; Hulet, A.; Chambers, J. Response of conifer-encroached shrublands in the Great Basin to prescribed fire and mechanical treatments. Rangel. Ecol. Manag. 2014, 67, 468–481. [Google Scholar] [CrossRef] [Green Version]
- Farzan, S.; Young, D.J.N.; Dedrick, A.G.; Hamilton, M.; Porse, E.C.; Coates, P.S.; Sampson, G. Western juniper management: Assessing strategies for improving greater sage-grouse habitat and rangeland productivity. Environ. Manag. 2015, 56, 675–683. [Google Scholar] [CrossRef] [Green Version]
- Gifford, G.F.; Shaw, C.B. Soil moisture patterns on two chained pinyon-juniper sites in Utah. J. Range Manag. 1973, 26, 436–440. [Google Scholar] [CrossRef]
- Everett, R.L.; Sharrow, S.H. Soil Water and Temperature in Harvested and Nonharvested Pinyon-Juniper Stands; Res. Paper INT-342; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1985; p. 5.
- Bates, J.D.; Miller, R.F.; Svejcar, T.J. Understory dynamics in cut and uncut western juniper woodlands. J. Range Manag. 2000, 53, 119–126. [Google Scholar] [CrossRef]
- Roundy, B.A.; Young, K.; Cline, N.; Hulet, A.; Miller, R.F.; Tausch, R.J.; Chambers, J.C.; Rau, B. Piñon-juniper reduction increases soil water availability of the resource growth pool. Rangel. Ecol. Manag. 2014, 67, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Young, K.R.; Roundy, B.A.; Eggett, D.L. Tree reduction and debris from mastication of Utah juniper alter the soil climate in sagebrush steppe. For. Ecol. Manag. 2013, 310, 777–785. [Google Scholar] [CrossRef] [Green Version]
- Tennesen, M. When juniper and woody plants invade, water may retreat. Science 2008, 322, 1630–1631. [Google Scholar] [CrossRef] [PubMed]
- Pyke, D.A.; Chambers, J.C.; Pellant, M.; Knick, S.T.; Miller, R.F.; Beck, J.L.; Doescher, P.S.; Schupp, E.W.; Roundy, B.A.; Brunson, M.; et al. Restoration Handbook for Sagebrush Steppe Ecosystems with Emphasis on Greater Sage-Grouse Habitat—Part. 1. Concepts for Understanding and Applying Restoration; Circular 1416; U.S. Department of the Interior, U.S. Geological Survey: Reston, VA, USA, 2015; p. 44. [CrossRef]
- Williams, C.J.; Pierson, F.B.; Kormos, P.R.; Al-Hamdan, O.Z.; Nouwakpo, S.K.; Weltz, M.A. Vegetation, hydrologic, and erosion responses of sagebrush steppe 9 yr following mechanical tree removal. Rangel. Ecol. Manag. 2019, 72, 47–68. [Google Scholar] [CrossRef]
- Fulbright, T.E.; Davies, K.W.; Archer, S.R. Wildlife responses to brush management: A contemporary evaluation. Rangel. Ecol. Manag. 2018, 71, 35–44. [Google Scholar] [CrossRef]
- Archer, S.R.; Davies, K.W.; Fulbright, T.E.; McDaniel, K.C.; Wilcox, B.P.; Predick, K.I. Brush management as a rangeland conservation strategy: A critical evaluation. In Conservation Benefits of Rangeland Practices: Assessment, Recommendations, and Knowledge Gaps; Briske, D.D., Ed.; Allen Press: Lawrence, KS, USA, 2011; pp. 105–170. [Google Scholar]
- Provencher, L.; Thompson, J. Vegetation responses to pinyon-juniper treatments in eastern Nevada. Rangel. Ecol. Manag. 2014, 67, 195–205. [Google Scholar] [CrossRef]
- Gottfried, G.J.; Severson, K.E. Managing pinyon-juniper woodlands. Rangelands 1994, 16, 234–236. [Google Scholar]
- Roccaforte, J.P.; Fulé, P.Z.; Covington, W.W. Monitoring landscape-scale ponderosa pine restoration treatment implementation and effectiveness. Restor. Ecol. 2010, 18, 820–833. [Google Scholar] [CrossRef]
- McIver, J.; Brunson, M. Multidisciplinary, multisite evaluation of alternative sagebrush steppe restoration treatments: The SageSTEP Project. Rangel. Ecol. Manag. 2014, 67, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Copeland, S.M.; Munson, S.M.; Bradford, J.B.; Butterfield, B.J.; Morgan, J. Influence of climate, post-treatment weather extremes, and soil factors on vegetation recovery after restoration treatments in the southwestern US. Appl. Veg. Sci. 2019, 22, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Benayas, J.M.R.; Newton, A.C.; Diaz, A.; Bullock, J.M. Enhancement of biodiversity and ecosystem services by ecological restoration: A meta-analysis. Science 2009, 325, 1121–1124. [Google Scholar] [CrossRef] [PubMed]
- Gurevitch, J.; Koricheva, J.; Nakagawa, S.; Stewart, G. Meta-analysis and the science of research synthesis. Nature 2018, 555, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Aparicio, L.; Lortie, C.J. Advancing plant ecology through meta-analyses. J. Ecol. 2014, 102, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Willms, J.; Bartuszevige, A.; Schwilk, D.W.; Kennedy, P.L. The effects of thinning and burning on understory vegetation in North America: A meta-analysis. For. Ecol. Manag. 2017, 392, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Shindler, B.; Gordon, R.; Brunson, M.W.; Olsen, C. Public perceptions of sagebrush ecosystem management in the Great Basin. Rangel. Ecol. Manag. 2011, 64, 335–343. [Google Scholar] [CrossRef]
- Redmond, M.D.; Zelikova, T.J.; Barger, N.N. Limits to understory plant restoration following fuel-reduction treatments in a pinon-juniper woodland. Environ. Manag. 2014, 54, 1139–1152. [Google Scholar] [CrossRef]
- West, N.E. Factors affecting treatment success in the pinyon-juniper type. In Proceedings: Second Utah Shrub Ecology Workshop; Johnson, K.L., Ed.; Utah State University: Logan, UT, USA, 1984; pp. 21–33. [Google Scholar]
- Williams, R.E.; Roundy, B.A.; Hulet, A.; Miller, R.F.; Tausch, R.J.; Chambers, J.C.; Matthews, J.; Schooley, R.; Eggett, D. Pretreatment tree dominance and conifer removal treatments affect plant succession in sagebrush communities. Rangel. Ecol. Manag. 2017, 70, 759–773. [Google Scholar] [CrossRef]
- Miller, R.F.; Chambers, J.C.; Pellant, M. A Field Guide for Selecting the Most Appropriate Treatment in Sagebrush and Piñon-Juniper Ecosystems in the Great Basin; Gen. Tech. Rep. RMRS-GTR-322; Station, R.M.R., Ed.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2014; p. 72.
- House, J.I.; Archer, S.; Breshears, D.D.; Scholes, R.J. Conundrums in mixed woody-herbaceous plant systems. J. Biogeogr. 2003, 30, 1763–1777. [Google Scholar] [CrossRef] [Green Version]
- Roundy, B.A.; Chambers, J.C.; Pyke, D.A.; Miller, R.F.; Tausch, F.J.; Schupp, E.W.; Rau, B.; Gruell, T. Resilience and resistance in sagebrush ecosystems are associated with seasonal soil temperature and water availability. Ecosphere 2018, 9, e02417. [Google Scholar] [CrossRef]
- Stevens, R. Restoration of native communities by chaining and seeding. In Proceedings: Ecology and Managment of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 285–289. [Google Scholar]
- Baughman, C.; Forbis, T.A.; Provencher, L. Response of two sagebrush sites to low-disturbance, mechanical removal of pinyon and juniper. Invasive Plant Sci. Manag. 2010, 3, 122–129. [Google Scholar] [CrossRef]
- Rosenstock, S.S.; Van Riper, C. Breeding bird responses to juniper woodland expansion. J. Range Manag. 2001, 54, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Bybee, J.; Roundy, B.A.; Young, K.R.; Hulet, A.; Roundy, D.B.; Crook, L.; Aanderud, Z.; Eggett, D.L.; Cline, N.L. Vegetation response to piñon and juniper tree shredding. Rangel. Ecol. Manag. 2016, 69, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Hessing, M.B.; Johnson, C.D.; Balda, R.P. Early secondary succession of a pinyon-juniper woodland in a northern Arizona powerline corridor. Southwest Nat. 1982, 27, 1–9. [Google Scholar] [CrossRef]
- Schott, M.R.; Pieper, R.D. Succssion in pinyon-juniper vegetation in New Mexico. Rangelands 1986, 8, 126–128. [Google Scholar]
- Urza, A.K.; Weisberg, P.J.; Chambers, J.C.; Dhaemers, J.M.; Board, D. Post-fire vegetation response at the woodland-shrubland interface is mediated by the pre-fire community. Ecosphere 2017, 8, e01851. [Google Scholar] [CrossRef]
- Bernau, C.R.; Strand, E.K.; Bunting, S.C. Fuel bed response to vegetation treatments in juniper-invaded sagebrush steppe. Fire Ecol. 2018, 14. [Google Scholar] [CrossRef]
- Huffman, D.W.; Stoddard, M.T.; Springer, J.D.; Crouse, J.E. Understory responses to tree thinning and seeding indicate stability of degraded pinyon-juniper woodlands. Rangel. Ecol. Manag. 2017, 70, 484–492. [Google Scholar] [CrossRef]
- Davenport, D.W.; Breshears, D.D.; Wilcox, B.P.; Allen, C.D. Viewpoint: Sustainability of pinon-juniper ecosystems—A unifying perspective of soil erosion thresholds. J. Range Manag. 1998, 51, 231–240. [Google Scholar] [CrossRef]
- Clary, W.P.; Jameson, D.A. Herbage production following tree and shrub removal in the pinyon-juniper type of Arizona. J. Range Manag. 1981, 34, 109–113. [Google Scholar] [CrossRef]
- Knoop, W.T.; Walker, B.H. Interactions of woody and herbaceous vegetation in a southern African savanna. J. Ecol. 1985, 73, 235–253. [Google Scholar] [CrossRef]
- Riginos, C. Grass competition suppresses tree growth across multiple demographic stages. Ecology 2009, 90, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riginos, C.; Grace, J.B.; Augustine, D.J.; Young, T.P. Local versus landscape-scale effects of savanna trees on grasses. J. Ecol. 2009, 97, 1337–1345. [Google Scholar] [CrossRef]
- Chambers, J.C.; Maestas, J.D.; Pyke, D.A.; Boyd, C.S.; Pellant, M.; Wuenschel, A. Using resilience and resistance concepts to manage persistent threats to sagebrush ecosystems and greater sage-grouse. Rangel. Ecol. Manag. 2017, 70, 149–164. [Google Scholar] [CrossRef]
- Barney, M.A.; Frischknecht, N.C. Vegetation changes following fire in the pinyon-juniper type of west-central Utah. J. Range Manag. 1974, 27, 91–96. [Google Scholar] [CrossRef]
- Kerns, B.K.; Day, M.A. The importance of disturbance by fire and other abiotic and biotic factors in driving cheatgrass invasion varies based on invasion stage. Biol. Invas. 2017, 19, 1853–1862. [Google Scholar] [CrossRef]
- Chambers, J.C.; Roundy, B.A.; Blank, R.R.; Meyer, S.E.; Whittaker, A. What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecol. Monogr. 2007, 77, 117–145. [Google Scholar] [CrossRef] [Green Version]
- Everett, R.L.; Sharrow, S.H. Understory response to tree harvesting of singleleaf pinyon and Utah juniper. Great Basin Nat. 1985, 45, 105–112. [Google Scholar]
- Bates, J.D.; Sharp, R.N.; Davies, K.W. Sagebrush steppe recovery after fire varies by development phase of Juniperus occidentalis woodland. Int. J. Wildl. Fire 2014, 23, 117. [Google Scholar] [CrossRef] [Green Version]
- Kerns, B.K.; Day, M.A. Fuel reduction, seeding, and vegetation in a juniper woodland. Rangel. Ecol. Manag. 2014, 67, 667–679. [Google Scholar] [CrossRef]
- Davies, K.W.; Rios, R.C.; Bates, J.D.; Johnson, D.D.; Kerby, J.; Boyd, C.S. To burn or not to burn: Comparing reintroducing fire with cutting an encroaching conifer for conservation of an imperiled shrub-steppe. Ecol. Evol. 2019, 9, 9137–9148. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.F.; Bates, J.D.; Svejcar, T.J.; Pierson, F.B.; Eddleman, L.E. Biology, Ecology, and Management of Western Juniper; Tech. Bull. 152; Oregon State University, Agricultural Experiment Station, Ed.; Oregon State University, Agricultural Experiment Station: Corvallis, OR, USA, 2005; p. 7. [Google Scholar]
- Skousen, J.G.; Davis, J.N.; Brotherson, J.D. Pinyon-juniper chaining and seeding for big game in central Utah. J. Range Manag. 1989, 42, 98–104. [Google Scholar] [CrossRef]
- Little, E.L. Managing southwestern piñon-juniper woodlands: The past half centrury and the future. In Managing Piñon-Juniper Ecosystems for Sustainability and Social Needs, 26–30 April 1993, Sante Fe, NM; Gen. Tech. Rep. RM-236; Aldon, E.F., Shaw, D.W., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1993; pp. 105–107. [Google Scholar]
- Tausch, R.J.; Miller, R.F.; Roundy, B.A.; Chambers, J.C. Piñon and Juniper Field Guide: Asking the Right Questions to Select Appropriate Management Actions; Circular 1335; U.S. Department of the Interior, U.S. Geological Survey: Reston, VA, USA, 2009; p. 96.
- Fairchild, J.A. Pinyon-juniper chaining design guidelines for big game wither range enhancement projects. In Proceedings: Ecology and Management of Pinyon-Juniper Communities within the Interior West, 15–18 September 1997, Provo, UT; RMRS-P-9; Monsen, S.B., Stevens, R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999; pp. 278–280. [Google Scholar]
- Crow, C.; Riper, C.V. Avian community responses to mechanical thinning of a pinyon-juniper woodland: Specialist sensitivity to tree reduction. Nat. Areas J. 2010, 30, 191–201. [Google Scholar] [CrossRef]
- Koniak, S. Broadcast Seeding Success in Eight Pinyon-Juniper Stands after Wildfire; Res. Note INT-334; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1983; p. 4.
- Huffman, D.W.; Stoddard, M.T.; Springer, J.D.; Crouse, J.E.; Chancellor, W.W. Understory plant community responses to hazardous fuels reduction treatments in pinyon-juniper woodlands of Arizona, USA. For. Ecol. Manag. 2013, 289, 478–488. [Google Scholar] [CrossRef]
- Ott, J.E.; McArthur, E.D.; Roundy, B.A. Vegetation of chained and non-chained seedings after wildfire in Utah. J. Range Manag. 2003, 56, 81–91. [Google Scholar] [CrossRef]
- Stevens, R. Thirty years of pinyon-juniper big game habitat improvement projects: What have we learned? In Proceedings—Pinyon-Juniper Conference; Gen. Tech. Rep. INT-215; Everett, R.L., Ed.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987; pp. 558–571. [Google Scholar]
- Stevens, R. Mechanical chaining and seeding. In Proceedings of the Ecology and Managment of Pinyon-Juniper Communities within the Interior West, Provo, UT, USA, 15–18 September 1997; Proc. RMRS-P-9. pp. 281–284. [Google Scholar]
- Daniel, T.W.; Rivers, R.J.; Isaacson, H.E.; Eberhard, E.J.; LeBaron, A.D. Management Alternatives for Pinyon-Juniper Woodlands. Part. A: The Ecology of the Pinyon-Juniper Type of the Colorado Plateau and the Basin and Range Provinces; Bureau of Land Management and Utah Agriculutural Experiment Station: Logan, UT, USA, 1966; p. 242. [Google Scholar]
- Tausch, R.J.; Tueller, P.T. Plant succession following chaining of pinyon-juniper woodlands in eastern Nevada. J. Range Manag. 1977, 30, 44–49. [Google Scholar] [CrossRef]
- Madsen, M.D.; Zvirzdin, D.L.; Petersen, S.L.; Hopkins, B.G.; Roundy, B.A. Anchor chaining’s influence on soil hydrology and seeding success in burned Pinon-Juniper woodlands. Rangel. Ecol. Manag. 2015, 68, 231–240. [Google Scholar] [CrossRef]
- Skousen, J.; Davis, J.N.; Brotherson, J.D. Comparison of vegetation patterns resulting from bulldozing and 2-way chaining on a Utah pinyon-juniper big game range. Great Basin Nat. 1986, 46, 508–512. [Google Scholar]
- Young, K.R.; Roundy, B.A.; Eggett, D.L. Plant establishment in masticated Utah juniper woodlands. Rangel. Ecol. Manag. 2013, 66, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Juran, C.; Roundy, B.A.; Davis, J.N. Wildfire rehabilitation success with and without chaining on the Henry Mountains, Utah. In Proceedings—Shrublands under Fire: Disturbance and Recovery in a Changing World, 6–8 June 2006, Cedar City, UT; RMRS-P-52; Kitchen, S.G., Pendleton, R.L., Monaco, T.A., Vernon, J., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2008; pp. 91–106. [Google Scholar]
- Thompson, T.W.; Roundy, B.A.; McArthur, E.D.; Jessop, B.D.; Waldron, B.; Davis, J.N. Fire rehabilitation using native and introduced species: A landscape trial. Rangel Ecol. Manag. 2006, 59, 237–248. [Google Scholar] [CrossRef]
- Clary, W.P. Plant Density and Cover Responses to Several Seeding Techniques Following Wildfire; Res. Note INT-384; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1988; p. 6.
- Stoddard, M.T.; Huffman, D.W.; Alcoze, T.M.; Fule, P.Z. Effects of slash on herbaceous communities in pinyon-juniper woodlands of northern Arizona. Rangel. Ecol. Manag. 2008, 61, 485–495. [Google Scholar] [CrossRef]
- Cline, N.L.; Roundy, B.A.; Pierson, F.B.; Kormos, P.; Williams, C.J. Hydrologic response to mechanical shredding in a juniper woodland. Rangel. Ecol. Manag. 2010, 63, 467–477. [Google Scholar] [CrossRef]
- Pierson, F.B.; Williams, C.J.; Kormos, P.R.; Al-Hamdan, O.Z. Short-term effects of tree removal on infiltration, runoff, and erosion in woodland-encroached sagebrush steppe. Rangel. Ecol. Manag. 2014, 67, 522–538. [Google Scholar] [CrossRef] [Green Version]
- Brockway, D.G.; Gatewood, R.G.; Paris, R.B. Restoring grassland savannas from degraded pinyon-juniper woodlands: Effects of mechanical overstory reduction and slash treatment alternatives. J. Environ. Manag. 2002, 64, 179–197. [Google Scholar] [CrossRef] [Green Version]
- Reiner, A.L.; Vaillant, N.M.; Fites-Kaufman, J.; Dailey, S.N. Mastication and prescribed fire impacts on fuels in a 25-year old ponderosa pine plantation, southern Sierra Nevada. For. Ecol. Manag. 2009, 258, 2365–2372. [Google Scholar] [CrossRef]
- Faist, A.; Stone, H.; Tripp, E. Impacts of mastication: Soil seed bank responses to a forest thinning treatment in three Colorado (USA) conifer forest types. Forests 2015, 6, 3060–3074. [Google Scholar] [CrossRef]
- Ross, M.R.; Castle, S.C.; Barger, N.N. Effects of fuels reductions on plant communities and soils in a piñon-juniper woodland. J. Arid Environ. 2012, 79, 84–92. [Google Scholar] [CrossRef]
- Miller, R.F.; Chambers, J.C.; Pyke, D.A.; Pierson, F.B.; Williams, J.C. A Review of Fire Effects on Vegetation and Soils in the Great Basin Region: Response and Ecological Site Characteristics; Gen. Tech. Rep. RMRS-GTR-308; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; p. 126.
- Murphy, G.E.P.; Romanuk, T.N. A meta-analysis of community response predictability to anthropogenic disturbances. Am. Nat. 2012, 180, 316–327. [Google Scholar] [CrossRef]
- Lavin, M.; Brummer, T.J.; Quire, R.; Maxwell, B.D.; Rew, L.J. Physical disturbance shapes vascular plant diversity more profoundly than fire in the sagebrush steppe of southeastern Idaho, USA. Ecol. Evol. 2013, 3, 1626–1641. [Google Scholar] [CrossRef]
- Pieper, R.D. Spatial variation of piñon-juniper woodlands in New Mexico. In Managing Pinyon-Juniper Ecosystems for Sustainability and Social Needs, 26–30 April 1993, Sante Fe, NM; Gen Tech. Rep. RM-236; Aldon, E.F., Shaw, D.W., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1993; pp. 89–92. [Google Scholar]
- West, N.E.; Tausch, R.J.; Tueller, P.T. A Management-Oriented Classification of Pinyon-Juniper Woodlands of the Great Basin; RMRS-GTR-12; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1998; p. 42.
- Leonard, S.G.; Miles, R.L.; Summerfield, H.A. Soils of the pinyon-juniper woodlands. In Proceedings—Pinyon-Juniper Conference, 13–16 January 1986, Reno, NV; Gen. Tech. Rep. INT-215; Everett, R.L., Ed.; U.S. Department of Agricuture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987; pp. 227–230. [Google Scholar]
- Evangelista, P.; Stohlgren, T.J.; Guenther, D.; Stewart, S. Vegetation response to fire and postburn seeding treatments in Juniper woodlands of the Grand Staircase-Escalante National Monument, Utah. West. N. Am. Nat. 2004, 64, 293–305. [Google Scholar]
- Pyke, D.A.; Shaff, S.E.; Lindgren, A.I.; Schupp, E.W.; Doescher, P.S.; Chambers, J.C.; Burnham, J.S.; Huso, M.M. Region-wide ecological responses of arid Wyoming big sagebrush communities to fuel treatments. Rangel. Ecol. Manag. 2014, 67, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Knutson, K.C.; Pyke, D.A.; Wirth, T.A.; Arkle, R.S.; Pilliod, D.S.; Brooks, M.L.; Chambers, J.C.; Grace, J.B. Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J. Appl Ecol. 2014, 51, 1414–1424. [Google Scholar] [CrossRef]
- Germino, M.J.; Barnard, D.M.; Davidson, B.E.; Arkle, R.S.; Pilliod, D.S.; Fisk, M.R.; Applestein, C. Thresholds and hotspots for shrub restoration following a heterogeneous megafire. Landsc. Ecol. 2018, 33, 1177–1194. [Google Scholar] [CrossRef]
- West, N.E.; Tausch, R.J.; Rea, K.H.; Tueller, P.T. Taxonomic determination, distribution, and ecological indicator values of sagebrush within pinyon-juniper woodlands of Great Basin. J. Range Manag. 1978, 31, 87–92. [Google Scholar] [CrossRef]
- Stevens, R. Species adapted for seeding mountain brush, big, black, and low sagebrush, and pinyon-juniper communities. In Managing Intermountain Rangelands—Improvement of Range and Wildlife Habitats: Proceedings, 15–17 September 1981, Twin Falls, ID, 22–24 June 1982, Elko, NV; Gen. Tech. Rep. INT-157; Monsen, S.B., Shaw, N., Eds.; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experimental Station: Ogden, UT, USA, 1983; pp. 78–82. [Google Scholar]
- Moir, W.H.; Carleton, J.O. Classification of pinyon-juniper (P-J) sites on national forests in the southwest. In Proceedings—Pinyon-Juniper Conference, 13–16 January 1986, Reno, NV; Gen. Tech. Rep. INT-215; Everett, R.L., Ed.; U.S. Department of Agricuture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987; pp. 216–226. [Google Scholar]
- West, N.E.; Rea, K.H.; Tausch, R.J. Basic synecological relationships in juniper-pinyon woodlands. In The Pinyon-juniper Ecosystem: A Symposium; Utah Agricultural Experiment Station: Logan, UT, USA, 1975; pp. 41–54. [Google Scholar]
- McArthur, E.D. Ecology, distribution, and values of sagebrush within the Intermountain Region. In Proceedings: Ecology And Management of Annual Rangelands, 18–21 May 1992, Boise, ID; Gen. Tech. Rep. INT-GTR-313; Monsen, S.B., Kitchen, S.G., Eds.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1994; pp. 347–351. [Google Scholar]
- Winward, A.H. Using sagebrush ecology in wildland management. In Proceedings of the First Utah Shrub Ecology Workshop; Johnson, K.L., Ed.; Utah State University, College of Natural Resources: Logan, UT, USA, 1983; pp. 15–19. [Google Scholar]
- West, N.E. (Ed.) Great Basin-Colorado Plateau Sagebrush Semi-Desert. In Temperate Deserts and Semi-Deserts; Elsevier: Amsterdam, The Netherlands, 1983; pp. 331–349. [Google Scholar]
- Wilder, L.E.; Veblen, K.E.; Schupp, E.W.; Monaco, T.A. Seedling emergence patterns of six restoration species in soils from two big sagebrush plant communities. West. N. Am. Nat. 2019, 79, 233–246. [Google Scholar] [CrossRef]
- Jensen, M.E. Interpretation of environmental gradients which influence sagebrush community distribution in northeastern Nevada. J. Range Manag. 1990, 43, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Thatcher, A.P. Distribution of sagebrush as related to site differences in Albany County, Wyoming. J. Range Manag. 1959, 12, 55–61. [Google Scholar] [CrossRef]
- Goodrich, S. Classification and capabilities of woody sagebrush communities of western North America with emphasis on sage-grouse habitat. In Sage-Grouse Habitat Restoration Symposium Proceedings, 4–7 June, Boise, ID; RMRS-P-38; Shaw, N.L., Pellant, M., Monsen, S.B., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005; pp. 17–37. [Google Scholar]
- Shultz, L.M. Monograph of Artemisia subgenus Tridentatae (Asteraceae-Anthemideae). Bot. Monogr. 2009, 89, 1–131. [Google Scholar]
- Wilder, L.E.; Veblen, K.E.; Gunnell, K.L.; Monaco, T.A. Influence of fire and mechanical sagebrush reduction treatments on restoration seedings in Utah, United States. Restor. Ecol. 2019, 27, 308–319. [Google Scholar] [CrossRef]
- Maestas, J.D.; Campbell, S.B. Mapping Potential Ecosystem Resilience and Resistance Across Sage-Grouse Range Using Soil Temperature and Moisture Regimes. 2016. Available online: http://www.usu.edu/saf/PJWoodlandsPositionStatement.pdf (accessed on 27 August 2020).
- Chambers, J.C.; Miller, R.F.; Board, D.I.; Pyke, D.A.; Roundy, B.A.; Grace, J.B.; Schupp, E.W.; Tausch, R.J. Resilience and resistance of sagebrush ecosystems: Implications for state and transition models and management treatments. Rangel. Ecol. Manag. 2014, 67, 440–454. [Google Scholar] [CrossRef] [Green Version]
- Riginos, C.; Veblen, K.E.; Thacker, E.T.; Gunnell, K.L.; Monaco, T.A. Disturbance type and sagebrush community type affect plant community structure after shrub reduction. Rangel. Ecol. Manag. 2019, 72, 619–631. [Google Scholar] [CrossRef]
- Chambers, J.C.; Bradley, B.A.; Brown, C.S.; D’Antonio, C.; Germino, M.J.; Grace, J.B.; Hardegree, S.P.; Miller, R.F.; Pyke, D.A. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 2014, 17, 360–375. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Pauchard, A.; Lenoir, J.; Nunez, M.A.; Geron, C.; Ven, A.; Bravo-Monasterio, P.; Teneb, E.; Nijs, I.; Milbau, A. Disturbance is the key to plant invasions in cold environments. Proc. Natl. Acad. Sci. USA 2016, 113, 14061–14066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, R.M.; Bakker, J.D.; Vincent, J.B.; Davies, G.M. Relative importance of abiotic, biotic, and disturbance drivers of plant community structure in the sagebrush steppe. Ecol. Appl. 2017, 27, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Urza, A.K.; Weisberg, P.J.; Chambers, J.C.; Board, D.; Flake, S.W. Seeding native species increases resistance to annual grass invasion following prescribed burning of semiarid woodlands. Biol. Invas. 2019, 21, 1993–2007. [Google Scholar] [CrossRef]
- Johnson, D.D.; Miller, R.F. Structure and development of expanding western juniper woodlands as influenced by two topographic variables. For. Ecol. Manag. 2006, 229, 7–15. [Google Scholar] [CrossRef]
- Jacobs, B.F.; Romme, W.H.; Allen, C.D. Mapping “old” vs. “young” pinon-juniper stands with a predictive topo-climatic model. Ecol. Appl. 2008, 18, 1627–1641. [Google Scholar] [CrossRef]
- Davis, J.N.; Harper, K.T. Weedy annuals and establishment of seeded species on a chained juniper-pinyon woodland in central Utah. In Wildland Shrub Dieoffs Following Excessivey Wet Periods: A Synthesis; McArthur, E.D., Romney, E.M., Smith, S.D., Tueller, P.T., Eds.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1990; pp. 72–79. [Google Scholar]
- Everett, R.L.; Ward, K. Early plant succession on pinyon-juniper controlled burns. Northwest Sci. 1984, 58, 57–68. [Google Scholar]
- James, J.J.; Davies, K.W.; Sheley, R.L.; Aanderud, Z.T. Linking nitrogen partitioning and species abundance to invasion resistance in the Great Basin. Oecologia 2008, 156, 637–648. [Google Scholar] [CrossRef]
- Leonard, E.D.; Monaco, T.A.; Stark, J.M.; Ryel, R.J. Invasive forb, annual grass, and exotic shrub competition with three sagebrush-steppe growth forms: Acquisition of a spring 15N tracer. Invasive Plant Sci. Manag. 2008, 1, 168–177. [Google Scholar] [CrossRef]
- Deines, L.; Rosentreter, R.; Eldridge, D.J.; Serpe, M.D. Germination and seedling establishment of two annual grasses on lichen-dominated biological soil crusts. Plant Soil 2007, 295, 23–35. [Google Scholar] [CrossRef]
- Cain, D. The Ely Chain: A Practical Handbook of Principles and Practices of Chaining and Vegetative Manipulation; U.S. Department of the Interior, Bureau of Land Management: Ely, NV, USA, 1971; p. 32.
- McKenzie, D.; Jensen, F.R.; Johnsen, T.N.; Young, J.A. Chains for mechanical brush control. Rangelands 1984, 6, 122–127. [Google Scholar]
- Evans, R.A.; Young, J.A. Plant succession following control of western juniper (Juniperus occidentalis) with picloram. Weed Sci. 1985, 33, 63–68. [Google Scholar] [CrossRef]
- Havrilla, C.A.; Faist, A.M.; Barger, N.N. Understory plant community responses to fuel-reduction treatments and seeding in an upland piñon-juniper woodland. Rangel. Ecol. Manag. 2017, 70, 609–620. [Google Scholar] [CrossRef]
- Stephens, G.J.; Johnston, D.B.; Jonas, J.L.; Paschke, M.W. Understory responses to mechanical treatment of pinyon-juniper in northwestern Colorado. Rangel. Ecol. Manag. 2016, 69, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Owen, S.M.; Sieg, C.H.; Gehring, C.A.; Bowker, M.A. Above- and belowground responses to tree thinning depend on the treatment of tree debris. For. Ecol. Manag. 2009, 259, 71–80. [Google Scholar] [CrossRef]
- Potts, J.B.; Stephens, S.L. Invasive and native plant responses to shrubland fuel reduction: Comparing prescribed fire, mastication, and treatment season. Biol. Conserv. 2009, 142, 1657–1664. [Google Scholar] [CrossRef]
- Coop, J.D.; Grant, T.A.; Magee, P.A.; Moore, E.A. Mastication treatment effects on vegetation and fuels in piñon-juniper woodlands of central Colorado, USA. For. Ecol. Manag. 2017, 396, 68–84. [Google Scholar] [CrossRef]
- Rubin, R.L.; Roybal, C.M. Plant community responses to mastication and mulching of one-seed juniper (Juniperus monosperma). Rangel. Ecol. Manag. 2018, 71, 753–756. [Google Scholar] [CrossRef]
- Young, K.R.; Roundy, B.R.; Eggett, D.L. Mechanical mastication of Utah juniper encroaching sagebrush steppe increases inorganic soil N. Appl. Environ. Soil Sci. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Aanderud, Z.T.; Schoolmaster, D.R.; Rigby, D.; Bybee, J.; Campbell, T.; Roundy, B.A. Soils mediate the impact of fine woody debris on invasive and native grasses as whole trees are mechanically shredded into firebreaks in piñon-juniper woodlands. J. Arid Environ. 2017, 137, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Koniak, S. Succession in pinyon-juniper woodlands following wildfire in the Great Basin. Great Basin Nat. 1985, 45, 556–566. [Google Scholar]
- Miller, R.F.; Chambers, J.C.; Pellant, M. A Field Guide for Rapid Assessment of Post-Wildfire Recovery Potential in Sagebrush and Pinon-Juniper Ecosystems in the Great Basin; Gen. Tech. Rep. RMRS-GTR-338; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2015; p. 70.
- Redmond, M.D.; Cobb, N.S.; Miller, M.E.; Barger, N.N. Long-term effects of chaining treatments on vegetation structure in pinon-juniper woodlands of the Colorado Plateau. For. Ecol. Manag. 2013, 305, 120–128. [Google Scholar] [CrossRef]
- Van Auken, O.W. Causes and consequences of woody plant encroachment into western North American grasslands. J. Environ. Manag. 2009, 90, 2931–2942. [Google Scholar] [CrossRef] [PubMed]
- Caracciolo, D.; Istanbulluoglu, E.; Noto, L.V. An ecohydrological cellular automata model investigation of juniper tree encroachment in a western North American landscape. Ecosystems 2017, 20, 1104–1123. [Google Scholar] [CrossRef]
- West, N.E. (Ed.) Overview of North American temperate deserts and semi-deserts. In Temperate Deserts and Semi-Deserts; Elsevier Scientific Publishing Company: Amsterdam, The Netherlands, 1983; pp. 321–330. [Google Scholar]
- Proctor, M.C.F.; Tuba, Z. Poikilohydry and homoihydry: Antithesis or spectrum of possibilities? New Phytol. 2002, 156, 327–349. [Google Scholar] [CrossRef] [Green Version]
- Hirsch-Schantz, M.C.; Monaco, T.A.; Call, C.A.; Sheley, R.L. Large-scale downy brome treatments alter plant-soil relationships and promote perennial grasses in salt desert shrublands. Rangel. Ecol. Manag. 2014, 67, 255–265. [Google Scholar] [CrossRef]
- Compagnoni, A.; Adler, P.B. Warming, soil moisture, and loss of snow increase Bromus tectorum’s population growth rate. Elementa 2014, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Condon, L.A.; Pyke, D.A. Filling the interspace-restoring arid land mosses: Source populations, organic matter, and overwintering govern success. Ecol. Evol. 2016, 6, 7623–7632. [Google Scholar] [CrossRef]
- Williams, C.J.; Snyder, K.A.; Pierson, F.B. Spatial and temporal variability of the impacts of pinyon and juniper reduction on hydrologic and rrosion processes across climatic gradients in the western US: A regional synthesis. Water 2018, 10, 1607. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.F.; Gatewood, R.G. Reintroduction of fire maintains structure of mechanically restored pinyon-juniper savanna (New Mexico). Ecol. Restor. 2002, 20, 207–208. [Google Scholar]
- Monaco, T.A.; Mangold, J.M.; Mealor, B.A.; Mealor, R.D.; Brown, C.S. Downy brome control and impacts on perennial grass abundance: A systematic review spanning 64 years. Rangel. Ecol. Manag. 2017, 70, 396–404. [Google Scholar] [CrossRef]
- Jacobs, B.F. Restoration of degraded transitional (piñon-juniper) woodland sites improves ecohydrologic condition and primes understory resilience to subsequent disturbance. Ecohydrology 2015, 8, 1417–1428. [Google Scholar] [CrossRef]
- Ashcroft, N.K.; Fernald, A.G.; VanLeeuwen, D.M.; Baker, T.T.; Cibils, A.F.; Boren, J.C. The effects of thinning trees and scattering slash on runoff and sediment yield within dense piñon-juniper woodlands in New Mexico, United States. J. Soil Water Conserv. 2017, 72, 122–130. [Google Scholar] [CrossRef]
- West, N.E.; Van Pelt, N.S. Successional patterns in pinyon-juniper woodlands. In Proceedings: Pinyon-Juniper Conference. USDA Forest Service, 13–16 January, Reno, NV; Gen. Tech. Rep. INT-215; Everett, R.L., Ed.; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987; pp. 43–52. [Google Scholar]
- Bates, J.D.; Boyd, C.S.; Davies, K.W. Longer-term post-fire succession on Wyoming big sagebrush steppe. Int. J. Wildl. Fire 2020, 29, 229–239. [Google Scholar] [CrossRef]
- Bates, J.D.; Davies, K.W. Seasonal burning of juniper woodlands and spatial recovery of herbaceous vegetation. For. Ecol. Manag. 2016, 361, 117–130. [Google Scholar] [CrossRef]
- Kane, J.M.; Meinhardt, K.A.; Chang, T.; Cardall, B.L.; Michalet, R.; Whitham, T.G. Drought-induced mortality of a foundation species (Juniperus monosperma) promotes positive afterlife effects in understory vegetation. Plant Ecol. 2011, 212, 733–741. [Google Scholar] [CrossRef]
- Coultrap, D.E.; Fulgham, K.E.; Lancaster, D.L.; Gustafson, J.; Lile, D.F.; George, M.R. Relationship between western Juniper (Juniperus occidentalis) and understory vegetation. Invasive Plant Sci. Manag. 2008, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Everett, R.L.; Sharrow, S.H. Response of Grass Species to Tree Harvesting in Singleleaf Pinyon-Utah Juniper Stands; Res. Paper INT-334; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1985; p. 7.
- Sheley, R.L.; Bates, J.D. Restoring western juniper- (Juniperus occidentalis) infested rangeland after prescribed fire. Weed Sci. 2008, 56, 469–476. [Google Scholar] [CrossRef]
- Davies, K.W.; Bates, J.D.; Madsen, M.D.; Nafus, A.M. Restoration of mountain big sagebrush steppe following prescribed burning to control western juniper. Environ. Manag. 2014, 53, 1015–1022. [Google Scholar] [CrossRef]
- Plummer, A.P.; Christensen, D.R.; Monsen, S.B. Restoring Big Game Range in Utah; Publ. 69-3.; Utah Division of Fish and Game: Salt Lake, UT, USA, 1969; p. 183. [Google Scholar]
- Redmond, M.D.; Golden, E.S.; Cobb, N.S.; Barger, N.N. Vegetation management across Colorado Plateau BLM Lands: 1950–2003. Rangel. Ecol. Manag. 2014, 67, 636–640. [Google Scholar] [CrossRef]
- Hourihan, E.; Schultz, B.W.; Perryman, B.L. Climatic influences on establishment pulses of four Artemisia species in Nevada. Rangel. Ecol. Manag. 2018, 71, 77–86. [Google Scholar] [CrossRef]
- Nelson, Z.J.; Weisberg, P.J.; Kitchen, S.G. Influence of climate and environment on post-fire recovery of mountain big sagebrush. Int. J. Wildl. Fire 2014, 23, 131–142. [Google Scholar] [CrossRef]
- Ziegenhagen, L.L.; Miller, R.F. Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. West. N. Am. Nat. 2009, 69, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Rosenstock, S.S.; Monsen, S.B.; Stevens, R.; Jorgensen, K.R. Mule Deer Diets on a Chained and Seeded Central Utah Pinyon-Juniper Range; Res. Paper INT-410; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1989; p. 4.
- Rosenstock, S.S.; Stevens, R. Herbivore effects on seeded alfalfa at four pinyon-juniper sites in central Utah. J. Range Manag. 1989, 42, 483–490. [Google Scholar] [CrossRef]
- Sorensen, G.E.; Kramer, D.W.; Cain, J.W.; Taylor, C.A.; Gipson, P.S.; Wallace, M.C.; Cox, R.D.; Ballard, W.B. Mule deer habitat selection following vegetation thinning treatments in New Mexico. Wildl. Soc. Bull. 2020, 44. [Google Scholar] [CrossRef]
- Gunnell, K.L.; Monaco, T.A.; Call, C.A.; Ransom, C.V. Seedling interference and niche differentiation between crested wheatgrass and contrasting native Great Basin species. Rangel. Ecol. Manag. 2010, 63, 443–449. [Google Scholar] [CrossRef]
- McAdoo, J.K.; Boyd, C.S.; Sheley, R.L. Site, competition, and plant stock influence transplant success of Wyoming big sagebrush. Rangel. Ecol. Manag. 2013, 66, 305–312. [Google Scholar] [CrossRef]
- Williams, J.R.; Morris, L.R.; Gunnell, K.L.; Johanson, J.K.; Monaco, T.A. Variation in sagebrush communities historically seeded with crested wheatgrass in the eastern Great Basin. Rangel. Ecol. Manag. 2017, 70, 683–690. [Google Scholar] [CrossRef]
- Rottler, C.M.; Burke, I.C.; Palmquist, K.A.; Bradford, J.B.; Lauenroth, W.K. Current reclamation practices after oil and gas development do not speed up succession or plant community recovery in big sagebrush ecosystems in Wyoming. Restor. Ecol. 2017, 26. [Google Scholar] [CrossRef]
- McIver, J.; Brunson, M.; Bunting, S.; Chambers, J.; Doescher, P.; Grace, J.; Hulet, A.; Johnson, D.; Knick, S.; Miller, R.; et al. A synopsis of short-term response to alternative restoration treatments in sagebrush-steppe: The SageSTEP Project. Rangel. Ecol. Manag. 2014, 67, 584–598. [Google Scholar] [CrossRef] [Green Version]
- Daryanto, S.; Wang, L.; Fu, B.; Zhao, W.; Wang, S. Vegetation responses and trade-offs with soil-related ecosystem services after shrub removal: A meta-analysis. Land Degrad. Dev. 2019, 30, 1219–1228. [Google Scholar] [CrossRef] [Green Version]
- Anadon, J.D.; Sala, O.E.; Turner, B.L., 2nd; Bennett, E.M. Effect of woody-plant encroachment on livestock production in North and South America. Proc. Natl. Acad. Sci. USA 2014, 111, 12948–12953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunson, M. Unwanted no more: Land use, ecosystem services, and opportunities for resilience in human-influenced shrublands. Rangelands 2014, 36, 5–11. [Google Scholar] [CrossRef]
- Archer, S.R.; Predick, K.I. An ecosystem services perspective on brush management: Research priorities for competing land-use objectives. J. Ecol. 2014, 102, 1394–1407. [Google Scholar] [CrossRef] [Green Version]
- Omernik, J.M.; Griffith, G.E. Ecoregions of the conterminous United States: Evolution of a hierarchical spatial framework. Environ. Manag. 2014, 54, 1249–1266. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.G. Ecoregions—The Ecosystem Geography of the Oceans and Continents; Springer: New York, NY, USA, 1995; p. 176. [Google Scholar]
- Benson, B. Technical Note: Pinyon and Utah Juniper Site Evaluation Procedure for Utah; U.S. Department of Agriculture, Natural Resources Conservation Service: Salt Lake, UT, USA, 2014; p. 7.
- Bonham, C.D. Measurements of Terrestrial Vegetation; Wiley-Blackwell: Chichester, UK, 2013. [Google Scholar]
- Bonham, C.C.; Mergen, D.E.; Montoya, S. Plant cover estimation: A contiguous Daubenmire frame. Rangelands 2004, 26, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J. Cryptogams, vascular plants, and soil hydrological relations: Some preliminary results from the semiarid woodlands of eastern Australia. Great Basin Nat. 1993, 53, 48–58. [Google Scholar]
- West, N.E. Structure and function of microphytic soil crusts in wildland ecosystems of arid and semi-arid regions. Adv. Ecol. Res. 1990, 20, 179–223. [Google Scholar]
- Canfield, R. Application of the line interception method in sampling range vegetation. J. For. 1941, 39, 388–394. [Google Scholar]
- Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 2010, 36, 1–48. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.L.; Oh, I.S.; Hayes, T.L. Fixed-versus random-effects models in meta-analysis: Model properties and an empirical comparison of differences in results. Br. J. Math. Stat. Psychol. 2009, 62, 97–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Koricheva, J.; Gurevitch, J. Uses and misuses of meta-analysis in plant ecology. J. Ecol. 2014, 102, 828–844. [Google Scholar] [CrossRef]
- Nakagawa, S.; Noble, D.W.; Senior, A.M.; Lagisz, M. Meta-evaluation of meta-analysis: Ten appraisal questions for biologists. BMC Biol. 2017, 15, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, S.; Cuthill, I.C. Effect size, confidence interval and statistical significance: A practical guide for biologists. Biol. Rev. 2007, 82, 591–605. [Google Scholar] [CrossRef]
- Curtis, P.S.; Wang, X. A meta-analysis of elevated CO2 effects on woody plant mass, form, and physiology. Oecologia 1998, 113, 299–313. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monaco, T.A.; Gunnell, K.L. Understory Vegetation Change Following Woodland Reduction Varies by Plant Community Type and Seeding Status: A Region-Wide Assessment of Ecological Benefits and Risks. Plants 2020, 9, 1113. https://doi.org/10.3390/plants9091113
Monaco TA, Gunnell KL. Understory Vegetation Change Following Woodland Reduction Varies by Plant Community Type and Seeding Status: A Region-Wide Assessment of Ecological Benefits and Risks. Plants. 2020; 9(9):1113. https://doi.org/10.3390/plants9091113
Chicago/Turabian StyleMonaco, Thomas A., and Kevin L. Gunnell. 2020. "Understory Vegetation Change Following Woodland Reduction Varies by Plant Community Type and Seeding Status: A Region-Wide Assessment of Ecological Benefits and Risks" Plants 9, no. 9: 1113. https://doi.org/10.3390/plants9091113