
Genomewide Analysis and Biological Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity from Three Bat Species

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Identification of 27 AMP Family Genes via the In Silico Analysis of R. ferrumequinum Genome
2.2. Differences in the Number of Functional Cathelicidin Genes among Seven Bat Species Belonging to Different Families
2.3. In Silico Determination of the Antimicrobial Activity Core Regions of Three Bat Cathelicidins
2.4. Confirmation of the Antibacterial Activity of Three Bat Cathelicidins
2.5. Confirmation of the Antifungal Activity of Three Bat Cathelicidins
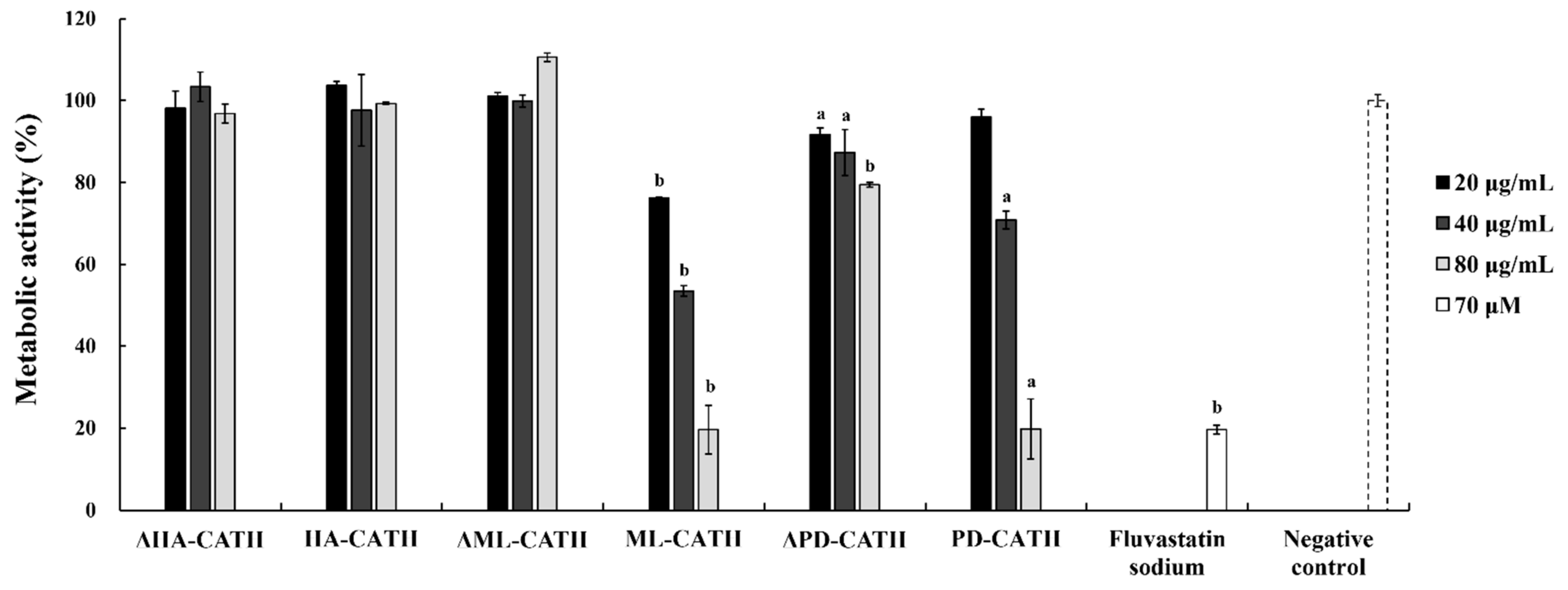
2.6. Negligible Cytotoxicity of Bat Cathelicidins against Human Keratinocytes
3. Discussion
4. Materials and Methods
4.1. In Silico Identification of Cathelicidin-like Sequences in the Genome of Seven Bat Species
4.2. In Silico Prediction of Putative Cathelicidins for Their Biological Activities
4.3. Peptide Synthesis
4.4. Evaluation of Antibacterial Activity
4.5. Evaluation of Antifungal Activity
4.6. In Vitro Cytotoxicity Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jones, K.E. Chiroptera (Bats). In eLS, 1st ed.; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Teeling, E.C.; Jones, G.; Rossiter, S.J. Phylogeny, Genes, and Hearing: Implications for the Evolution of Echolocation in Bats. In Bat Bioacoustics; Fenton, M.B., Grinnell, A.D., Popper, A.N., Fay, R.R., Eds.; Springer: New York, NY, USA, 2016; pp. 25–54. [Google Scholar]
- Prat, Y.; Azoulay, L.; Dor, R.; Yovel, Y. Crowd vocal learning induces vocal dialects in bats: Playback of conspecifics shapes fundamental frequency usage by pups. PLoS Biol. 2017, 15, e2002556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, N.B. An Eocene Big Bang for Bats. Science 2005, 307, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Jebb, D.; Teeling, E.C. Blood miRNomes and transcriptomes reveal novel longevity mechanisms in the long-lived bat, Myotis myotis. BMC Genom. 2016, 17, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E.; et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Jebb, D.; Huang, Z.; Pippel, M.; Hughes, G.M.; Lavrichenko, K.; Devanna, P.; Winkler, S.; Jermiin, L.S.; Skirmuntt, E.C.; Katzourakis, A.; et al. Six reference-quality genomes reveal evolution of bat adaptations. Nature 2020, 583, 578–584. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Matin, M.M.; Uzzaman, M.; Chowdhury, S.A.; Bhuiyan, M.M.H. In vitro antimicrobial, physicochemical, pharmacokinetics and molecular docking studies of benzoyl uridine esters against SARS-CoV-2 main protease. J. Biomol. Struct. Dyn. 2022, 40, 3668–3680. [Google Scholar] [CrossRef]
- Yang, H.; Rao, Z. Structural biology of SARS-CoV-2 and implications for therapeutic development. Nat. Rev. Microbiol. 2021, 19, 685–700. [Google Scholar] [CrossRef]
- Wang, L.-F.; Gamage, A.M.; Chan, W.O.Y.; Hiller, M.; Teeling, E.C. Decoding bat immunity: The need for a coordinated research approach. Nat. Rev. Immunol. 2021, 21, 269–271. [Google Scholar] [CrossRef]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef]
- Hayman, D.T.S. Bat tolerance to viral infections. Nat. Microbiol. 2019, 4, 728–729. [Google Scholar] [CrossRef]
- Bhattacharyya, S. Inflammation During Virus Infection: Swings and Roundabouts. In Dynamics of Immune Activation in Viral Diseases; Bramhachari, P.V., Ed.; Springer: Singapore, 2020; pp. 43–59. [Google Scholar]
- Teeling, E.; Vernes, S.C.; Davalos, L.M.; Ray, D.A.; Gilbert, M.T.P.; Myers, E.; Bat1K Consortium. Bat biology, genomes, and the Bat1K project: To generate chromosome-level genomes for all living bat species. Annu. Rev. Anim. Biosci. 2018, 6, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K.A.; Fang, X.; Wynne, J.W.; Xiong, Z.; Baker, M.L.; Zhao, W.; et al. Comparative Analysis of Bat Genomes Provides Insight into the Evolution of Flight and Immunity. Science 2013, 339, 456–460. [Google Scholar] [CrossRef] [Green Version]
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef]
- Zhan, L.-J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Nizet, V.; Ohtake, T.; Lauth, X.; Trowbridge, J.; Rudisill, J.; Dorschner, R.A.; Pestonjamasp, V.; Piraino, J.; Huttner, K.; Gallo, R.L. Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature 2001, 414, 454–457. [Google Scholar] [CrossRef]
- Mor, A. Multifunctional host defense peptides: Antiparasitic activities. FEBS J. 2009, 276, 6474–6482. [Google Scholar] [CrossRef]
- De Ullivarri, M.F.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell. Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef]
- Carriel-Gomes, M.C.; Kratz, J.M.; Barracco, M.A.; Bachére, E.; Barardi, C.R.M.; Simões, C.M.O. In vitro antiviral activity of antimicrobial peptides against herpes simplex virus 1, adenovirus, and rotavirus. Mem. Inst. Oswaldo Cruz 2007, 102, 469–472. [Google Scholar] [CrossRef]
- Cho, H.S.; Yum, J.; Larivière, A.; Lévêque, N.; Le, Q.V.C.; Ahn, B.; Jeon, H.; Hong, K.; Soundrarajan, N.; Kim, J.H.; et al. Opossum Cathelicidins Exhibit Antimicrobial Activity Against a Broad Spectrum of Pathogens Including West Nile Virus. Front. Immunol. 2020, 11, 347. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, P.E.; McHugh, B.; Findlay, E.G.; Mackellar, A.; MacKenzie, K.J.; Gallo, R.L.; Govan, J.R.W.; Simpson, A.J.; Davidson, D.J. Cathelicidin Host Defence Peptide Augments Clearance of Pulmonary Pseudomonas aeruginosa Infection by Its Influence on Neutrophil Function In Vivo. PLoS ONE 2014, 9, e99029. [Google Scholar] [CrossRef]
- Choi, K.-Y.G.; Napper, S.; Mookherjee, N. Human cathelicidin LL-37 and its derivative IG-19 regulate interleukin-32-induced inflammation. Immunology 2014, 143, 68–80. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial Peptides Human β-Defensins Stimulate Epidermal Keratinocyte Migration, Proliferation and Production of Proinflammatory Cytokines and Chemokines. J. Investig. Dermatol. 2007, 127, 594–604. [Google Scholar] [CrossRef] [Green Version]
- Tomasinsig, L.; Zanetti, M. The Cathelicidins-Structure, Function and Evolution. Curr. Protein Pept. Sci. 2005, 6, 23–34. [Google Scholar] [CrossRef]
- Sørensen, O.E.; Follin, P.; Johnsen, A.H.; Calafat, J.; Tjabringa, G.S.; Hiemstra, P.; Borregaard, N. Human cathelicidin, hCAP-18, is processed to the antimicrobial peptide LL-37 by extracellular cleavage with proteinase 3. Blood 2001, 97, 3951–3959. [Google Scholar] [CrossRef] [Green Version]
- Gudmundsson, G.H.; Agerberth, B.; Odeberg, J.; Bergman, T.; Olsson, B.; Salcedo, R. The Human Gene FALL39 and Processing of the Cathelin Precursor to the Antibacterial Peptide LL-37 in Granulocytes. Eur. J. Biochem. 1996, 238, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Van Harten, R.M.; Van Woudenbergh, E.; Van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory Antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasundarapandian, S.; Cho, H.-S.; Prathap, S.; Kang, M.; Choi, M.; Lee, Y.; Jeon, H.; Song, H.; Kim, J.-H.; Park, C. Cathelicidin ΔPb-CATH4 derived from Python bivittatus accelerates the healing of Staphylococcus aureus-infected wounds in mice. Amino Acids 2021, 53, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Cho, Y.; Dinh, N.-N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a Cathelin-Associated Antimicrobial Peptide of Human Neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [CrossRef] [Green Version]
- Gallo, R.L.; Kim, K.J.; Bernfield, M.; Kozak, C.A.; Zanetti, M.; Merluzzi, L.; Gennaro, R. Identification of CRAMP, a Cathelin-related Antimicrobial Peptide Expressed in the Embryonic and Adult Mouse. J. Biol. Chem. 1997, 272, 13088–13093. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Enée, E.; Andre-Vallee, C.; Falcone, M.; Sun, J.; Diana, J. Intestinal Cathelicidin Antimicrobial Peptide Shapes a Protective Neonatal Gut Microbiota Against Pancreatic Autoimmunity. Gastroenterology 2022, 162, 1288–1302.e16. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, J.; Qin, Z.; Dong, C.; Yang, H.; Sun, J.; Xu, W.; Wei, L. Endogenous cathelicidin is required for protection against ZIKV-caused testis damage via inactivating virons. Antivir. Res. 2022, 198, 105248. [Google Scholar] [CrossRef]
- Rather, I.A.; Sabir, J.S.M.; Asseri, A.H.; Ali, S. Antifungal Activity of Human Cathelicidin LL-37, a Membrane Disrupting Peptide, by Triggering Oxidative Stress and Cell Cycle Arrest in Candida auris. J. Fungi 2022, 8, 204. [Google Scholar] [CrossRef]
- Feng, G.; Wei, L.; Che, H.; Shen, Y.; Mi, K.; Bian, H.; Yang, H.; Wu, J.; Mu, L. Cathelicidin-NV from Nanorana ventripunctata effectively protects HaCaT cells, ameliorating ultraviolet B-induced skin photoaging. Peptides 2022, 150, 170712. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, F.; Guo, Z.; Chen, Y.; Zhang, M.; Yu, H.; Wang, Y. Characterization of a Cathelicidin from the Colubrinae Snake, Sinonatrix annularis. Zool. Sci. 2019, 36, 68–76. [Google Scholar] [CrossRef]
- Kalita, D.J. Characterization of cathelicidin gene from buffalo (Bubalus bubalis). Afr. J. Biotechnol. 2015, 14, 758–763. [Google Scholar]
- Dalla Valle, L.; Benato, F.; Paccanaro, M.C.; Alibardi, L. Bioinformatic and molecular characterization of cathelicidin-like peptides isolated from the green lizard Anolis carolinensis (Reptilia: Lepidosauria: Iguanidae). Ital. J. Zool. 2013, 80, 177–186. [Google Scholar] [CrossRef]
- De la Lastra, J.M.P.; Asensio-Calavia, P.; González-Acosta, S.; Baca-González, V.; Morales-delaNuez, A. Bioinformatic Analysis of Genome-Predicted Bat Cathelicidins. Molecules 2021, 26, 1811. [Google Scholar] [CrossRef]
- Belov, K.; Sanderson, C.E.; Deakin, J.E.; Wong, E.S.; Assange, D.; McColl, K.A.; Gout, A.; de Bono, B.; Barrow, A.D.; Speed, T.P.; et al. Characterization of the opossum immune genome provides insights into the evolution of the mammalian immune system. Genome Res. 2007, 17, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.K.; Le, M.T.; Nguyen, D.T.; Choi, H.; Kim, W.; Kim, J.H.; Chun, J.; Hyeon, J.; Seo, K.; Park, C. Genome-level identification, gene expression, and comparative analysis of porcine β-defensin genes. BMC Genet. 2012, 13, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.; Ahn, B.; Yum, J.; Cho, H.; Choi, M.; Hong, K.; Choi, Y.; Kim, J.; Park, C. Influence of habitat change from land to sea on the evolution of antimicrobial peptide gene families, including β-defensin gene clusters, in mammals. J. Zool. Syst. Evol. Res. 2021, 59, 510–521. [Google Scholar] [CrossRef]
- Cho, H.S.; Soundrarajan, N.; Le Van Chanh, Q.; Jeon, H.; Cha, S.Y.; Kang, M.; Ahn, B.; Hong, K.; Song, H.; Kim, J.H.; et al. The novel cathelicidin of naked mole rats, Hg-CATH, showed potent antimicrobial activity and low cytotoxicity. Gene 2018, 676, 164–170. [Google Scholar] [CrossRef]
- Ahn, B.; Jeon, H.; Cho H-s Nagasundarapandian, S.; Park, C. Sequence polymorphisms of PR39 cathelicidins and extensive copy variations in commercial pig breeds. Gene 2022, 822, 146323. [Google Scholar] [CrossRef]
- Zanetti, M. Cathelicidins, multifunctional peptides of the innate immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef]
- Pérez de Lastra, J.M.; Garrido-Orduña, C.; Borges, L.; Tejera, D.; Marino, M.; González-Guerra, E.; Borges, A.A. Antimicrobial activity of cathelicidins of mammals from avian, aquatic and terrestrial environments. In Proceedings of the IV International Conference on Antimicrobial Research ICAR, Torremolinos, Málaga, Spain, 29 June–1 July 2016. [Google Scholar]
- Taylor, S.W.; Craig, A.G.; Fischer, W.H.; Park, M.; Lehrer, R.I. Styelin D, an Extensively Modified Antimicrobial Peptide from Ascidian Hemocytes. J. Biol. Chem. 2000, 275, 38417–38426. [Google Scholar] [CrossRef] [Green Version]
- Dennison, S.R.; Harris, F.; Bhatt, T.; Singh, J.; Phoenix, D.A. The effect of C-terminal amidation on the efficacy and selectivity of antimicrobial and anticancer peptides. Mol. Cell Biochem. 2009, 332, 43. [Google Scholar] [CrossRef] [Green Version]
- Simmaco, M.; Kreil, G.; Barra, D. Bombinins, antimicrobial peptides from Bombina species. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1551–1555. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Soundrarajan, N.; Lee, J.; Cho, H.-S.; Choi, M.; Cha, S.-Y.; Ahn, B.; Jeon, H.; Le, M.T.; Song, H.; et al. Genomewide Analysis of the Antimicrobial Peptides in Python bivittatus and Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity. Antimicrob. Agents Chemother. 2017, 61, e00530-17. [Google Scholar] [CrossRef] [Green Version]
- Nagant, C.; Pitts, B.; Nazmi, K.; Vandenbranden, M.; Bolscher, J.G.; Stewart, P.S.; Dehaye, J.-P. Identification of Peptides Derived from the Human Antimicrobial Peptide LL-37 Active against Biofilms Formed by Pseudomonas aeruginosa Using a Library of Truncated Fragments. Antimicrob. Agents Chemother. 2012, 56, 5698–5708. [Google Scholar] [CrossRef] [Green Version]
- Travis, S.M.; Anderson, N.N.; Forsyth, W.R.; Espiritu, C.; Conway, B.D.; Greenberg, E.P.; McCray, P.; Lehrer, R.I.; Welsh, M.J.; Tack, B.F. Bactericidal Activity of Mammalian Cathelicidin-Derived Peptides. Infect. Immun. 2000, 68, 2748–2755. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.; Molhoek, E.M.; Veldhuizen, E.J.A.; Bokhoven, J.L.M.T.v.; Wagendorp, E.; Bikker, F.; Haagsman, H.P. Identification of chicken cathelicidin-2 core elements involved in antibacterial and immunomodulatory activities. Mol. Immunol. 2009, 46, 2465–2473. [Google Scholar] [CrossRef]
- Hertog, A.L.D.; Marle, J.V.; Veerman, E.C.I.; Valentijn-Benz, M.; Nazmi, K.; Kalay, H.; Grün, C.H.; Van’t Hof, W.; Bolscher, J.G.; Nieuw Amerongen, A.V. The human cathelicidin peptide LL-37 and truncated variants induce segregation of lipids and proteins in the plasma membrane of Candida albicans. Biol. Chem. 2006, 387, 1495–1502. [Google Scholar] [CrossRef]
- Taheri, B.; Mohammadi, M.; Nabipour, I.; Momenzadeh, N.; Roozbehani, M. Identification of novel antimicrobial peptide from Asian sea bass (Lates calcarifer) by in silico and activity characterization. PLoS ONE 2018, 13, e0206578. [Google Scholar] [CrossRef]
- Peel, E.; Cheng, Y.; Djordjevic, J.T.; Fox, S.; Sorrell, T.C.; Belov, K. Cathelicidins in the Tasmanian devil (Sarcophilus harrisii). Sci. Rep. 2016, 6, 35019. [Google Scholar] [CrossRef] [Green Version]
- Soundrarajan, N.; Cho, H.-S.; Ahn, B.; Choi, M.; Thong, L.M.; Choi, H.; Cha, S.-Y.; Kim, J.-H.; Park, C.-K.; Seo, K.; et al. Green fluorescent protein as a scaffold for high efficiency production of functional bacteriotoxic proteins in Escherichia coli. Sci. Rep. 2016, 6, 20661. [Google Scholar] [CrossRef] [Green Version]
- Arqué, X.; Torres, M.D.T.; Patiño, T.; Boaro, A.; Sánchez, S.; de la Fuente-Nunez, C. Autonomous Treatment of Bacterial Infections in Vivo Using Antimicrobial Micro- and Nanomotors. ACS Nano 2022, 16, 7547–7558. [Google Scholar] [CrossRef]
- Brandelli, A. Nanostructures as Promising Tools for Delivery of Antimicrobial Peptides. Mini Rev. Med. Chem. 2012, 12, 731–741. [Google Scholar] [CrossRef]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M. The role of cathelicidins in the innate host defenses of mammals. Curr. Issues Mol. Biol. 2005, 7, 179–196. [Google Scholar]
- Jones, K.E.; Purvis, A.; MacLarnon, A.N.N.; Bininda-Emonds, O.R.P.; Simmons, N.B. A phylogenetic supertree of the bats (Mammalia: Chiroptera). Biol. Rev. 2002, 77, 223–259. [Google Scholar] [CrossRef]
- Barlow, P.G.; Svoboda, P.; Mackellar, A.; Nash, A.A.; York, I.A.; Pohl, J.; Davidson, D.J.; Donis, R.O. Antiviral Activity and Increased Host Defense against Influenza Infection Elicited by the Human Cathelicidin LL-37. PLoS ONE 2011, 6, e25333. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Q.; Bhattacharya, S.; Waheed, A.A.; Tong, X.; Hong, A.; Heck, S.; Curreli, F.; Goger, M.; Cowburn, D.; et al. A Cell-penetrating Helical Peptide as a Potential HIV-1 Inhibitor. J. Mol. Biol. 2008, 378, 565–580. [Google Scholar] [CrossRef] [Green Version]
- Ben Mechlia, M.; Belaid, A.; Castel, G.; Jallet, C.; Mansfield, K.L.; Fooks, A.R.; Hani, K.; Tordo, N. Dermaseptins as potential antirabies compounds. Vaccine 2019, 37, 4694–4700. [Google Scholar] [CrossRef]
- Damour, A.; Garcia, M.; Cho, H.-S.; Larivière, A.; Lévêque, N.; Park, C.; Bodet, C. Characterisation of Antiviral Activity of Cathelicidins from Naked Mole Rat and Python bivittatus on Human Herpes Simplex Virus 1. Pharmaceuticals 2021, 14, 715. [Google Scholar] [CrossRef]
- Enayathullah, M.G.; Parekh, Y.; Banu, S.; Ram, S.; Nagaraj, R.; Kumar, B.K.; Idris, M.M. Gramicidin S and melittin: Potential anti-viral therapeutic peptides to treat SARS-CoV-2 infection. Sci. Rep. 2022, 12, 3446. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Lévêque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K3]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bairoch, A. UniProtKB/Swiss-Prot. In Plant Bioinformatics: Methods and Protocols; Edwards, D., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 89–112. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic. Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Torrent, M.; Nogués, V.M.; Boix, E. A theoretical approach to spot active regions in antimicrobial proteins. BMC Bioinform. 2009, 10, 373. [Google Scholar] [CrossRef] [Green Version]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED protein structure prediction server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Cantón, E.; Espinel-Ingroff, A.; Pemán, J. Trends in antifungal susceptibility testing using CLSI reference and commercial methods. Expert Rev. Anti-Infect. Ther. 2009, 7, 107–119. [Google Scholar] [CrossRef]
- Ballard, E.; Yucel, R.; Melchers, W.J.G.; Brown, A.J.P.; Verweij, P.E.; Warris, A. Antifungal Activity of Antimicrobial Peptides and Proteins against Aspergillus fumigatus. J. Fungi 2020, 6, 65. [Google Scholar] [CrossRef]


{kind=link}
| Bat Cathelicidin-Derived Peptides | Sequence | Length | <H> a | zb (+) | Molecular Weight (Da) | Similarity | ||
|---|---|---|---|---|---|---|---|---|
| AMP c | Source | (%) d | ||||||
| ΔHA-CATH | LLRRGGRKIGQGLERIGQRIQGF | 23 | 31 | 5 | 2609.08 | SMAP-29 | Ovis aries | 46.87 |
| HA-CATH | ILGRLRDLLRRGGRKIGQGLERIGQRIQGFFSNREPMEES | 40 | 30 | 4 | 4640.37 | K9CATH | Canis familiaris | 51.22 |
| ΔML-CATH | GIFILKHRRPIGRGIEIT | 18 | 50 | 3 | 2076.52 | Temporin-CPb | Lithobates capito | 40 |
| ML-CATH | LNPLIKAGIFILKHRRPIGRGIEITGRGIKKFFSK | 35 | 40 | 8 | 3975.87 | Palustrin-2CG1 | Amolops chunganensis | 38.64 |
| ΔPD-CATH | IAGRIAGKLIGDAINRHRERNRQRR | 25 | 36 | 6 | 2927.37 | TP4 | Oreochromis niloticus | 44.83 |
| PD-CATH | ILGPALRIGGRIAGRIAGKLIGDAINRHRERNRQRRG | 37 | 35 | 8 | 4088.79 | HKPLP | Hippocampus kuda | 43.59 |
| Strain | Minimum Inhibitory Concentration (μg/mL, μM) | ||||||
|---|---|---|---|---|---|---|---|
| HA-CATH | ML-CATH | PD-CATH | chloramphenicol | Ampicillin | Gentamicin | ||
| Gram-negative bacteria | Escherichia coli ATCC 25922 | 18 (3.9) | 2 (0.5) | 7 (1.7) | 3 (9.3) | 5 (14.3) | 1 (2.1) |
| Pseudomonas aeruginosa ATCC 27853 | >40 (8.6) | >40 (10.1) | >40 (9.8) | 80 (247.6) | >640 (1831.7) | 1 (2.1) | |
| Salmonella enterica serovar Typhimurium ATCC 14028 | >40 (8.6) | 21 (5.3) | 22 (5.4) | 5 (15.5) | >80 (228.8) | 1 (2.1) | |
| Acinetobacter baumannii KCTC 23254 | 5 (1.1) | 4 (1.0) | 4 (1.0) | 38 (117.6) | >80 (228.8) | 10 (21.0) | |
| Klebsiella pneumoniae subsp. pneumoniae KCTC 1726 | 34 (7.3) | 12 (3.0) | 13 (3.2) | > 80 (247.6) | >80 (228.8) | 19 (39.80 | |
| Enterobacter cloacae subsp. cloacae ATCC 13047 | >40 (8.6) | 35 (8.8) | 30 (7.3) | 5 (15.5) | >80 (228.8) | 1 (2.1) | |
| Gram-positive bacteria | Staphylococcus aureus ATCC 6538 | 26 (5.6) | 1 (0.3) | 3 (0.7) | 7.5 (23.2) | 2 (5.7) | 1 (2.1) |
| Bacillus cereus ATCC 10876 | 25 (5.4) | 3 (0.8) | 6 (1.5) | 10 (31.0) | 80 (228.8) | 1 (2.1) | |
| Enterococcus faecalis ATCC 29212 | >40 (8.6) | 5 (1.3) | 6 (1.5) | 10 (31.0) | 10 (28.6) | 90 (189.0) | |
| Streptococcus agalactiae ATCC 27956 | >40 (8.6) | 8 (2.0) | 8 (2.0) | 5 (15.5) | 4 (11.4) | 75 (157.5) | |
| Streptococcus dysgalactiae ATCC 27957 | >40 (8.6) | 14 (3.5) | 18 (4.4) | 4 (12.4) | 2 (5.7) | 15 (31.5) | |
| Streptococcus equi subsp. zooepidemicus ATCC 43079 | >40 (8.6) | 9 (2.3) | 17 (4.2) | 5 (15.5) | 2 (5.7) | 45 (94.5) | |
| Strain | MIC (μg/mL, μM) | ||||||
|---|---|---|---|---|---|---|---|
| ΔHA-CATH | HA-CATH | ΔML-CATH | ML-CATH | ΔPD-CATH | PD-CATH | Ciclopirox a | |
| Candida albicans KCTC 7270 | >40 (15.3) | >40 (8.6) | >40 (19.3) | 40 (10.1) | 25 (8.5) | 5 (1.2) | 3.5 (16. 9) |
| Cryptococcus cuniculi KCTC 17232 | >40 (15.3) | 45 (9.7) | >40 (19.3) | 5 (1.3) | 15 (5.1) | 5 (1.2) | 0.5 (2.4) |
| Treatment | Concentration (μg/mL) | Cell Viability ± SD (%) |
|---|---|---|
| ΔHA-CATH | 64 | 100.5 ± 0.5 |
| 160 | 97.7 ± 4.3 | |
| HA-CATH | 64 | 100 ± 0.3 |
| 160 | 97.5 ± 0.6 | |
| ΔML-CATH | 64 | 100.6 ± 1.3 |
| 160 | 96.0 ± 0.8 | |
| ML-CATH | 64 | 99.7 ± 0.8 |
| 160 | 97 ± 1.8 | |
| ΔPD-CATH | 64 | 100.3 ± 0.3 |
| 160 | 98.4 ± 2.3 | |
| PD-CATH | 64 | 100.5 ± 0.2 |
| 160 | 10.4 ± 0.4 * | |
| Melittin | 64 | 7.2 ± 0.03 * |
| Triton X-100 | - | 8.9 ± 0.05 * |
| Negative control | - | 99.5 ± 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.; Cho, H.-s.; Ahn, B.; Prathap, S.; Nagasundarapandian, S.; Park, C. Genomewide Analysis and Biological Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity from Three Bat Species. Antibiotics 2022, 11, 989. https://doi.org/10.3390/antibiotics11080989
Choi M, Cho H-s, Ahn B, Prathap S, Nagasundarapandian S, Park C. Genomewide Analysis and Biological Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity from Three Bat Species. Antibiotics. 2022; 11(8):989. https://doi.org/10.3390/antibiotics11080989
Chicago/Turabian StyleChoi, Munjeong, Hye-sun Cho, Byeongyong Ahn, Somasundaram Prathap, Soundrarajan Nagasundarapandian, and Chankyu Park. 2022. "Genomewide Analysis and Biological Characterization of Cathelicidins with Potent Antimicrobial Activity and Low Cytotoxicity from Three Bat Species" Antibiotics 11, no. 8: 989. https://doi.org/10.3390/antibiotics11080989